29.7 Þróun prímata
29.7 Þróun prímata
Markmið náms
Í lok þessa hluta muntu geta gert eftirfarandi:
Ættbálkur prímata (Primates) í flokki spendýra (Mammalia) inniheldur lemúra, tarsíera, apa, mannapa og menn. Prímatar aðrir en menn lifa fyrst og fremst á hitabeltis- eða heittempruðum svæðum Suður-Ameríku, Afríku og Asíu. Þeir eru breytilegir að stærð, allt frá múslemúrnum sem vegur 30 grömm upp í fjallagórilluna sem vegur 200 kílógrömm. Einkenni og þróun prímata vekja sérstakan áhuga okkar þar sem þau gera okkur kleift að skilja þróun okkar eigin tegundar.
Einkenni prímata
Allar tegundir prímata búa yfir aðlögunum til að klifra í trjám, því þær eru komnar af trjábúum. Þessi trjáarfur prímata hefur leitt til handa og fóta sem eru aðlagaðir klifri eða armasveiflu, það er að sveifla sér milli trjáa með handleggjunum. Þessar aðlaganir fela meðal annars í sér snúningslið í öxl, stórutá sem er vel aðskilin frá hinum tánum (nema hjá mönnum), þumal sem er nægilega aðskilinn frá fingrum til að grípa um greinar og þrívíddarsjón, þar sem sjónsvið augnanna skarast og gera dýptarskynjun og fjarlægðarmat möguleg. Önnur einkenni prímata eru heilar sem eru stærri en hjá flestum öðrum spendýrum, klær sem hafa breyst í flatar neglur, yfirleitt aðeins eitt afkvæmi í hverri meðgöngu og tilhneiging til uppréttrar líkamsstöðu.
Ættbálki prímata er skipt í tvo hópa: Strepsirrhini („sveigð nef“) og Haplorhini („einföld nef“). Strepsirrhini, einnig kallaðir votnefsprímatar, innihalda hálfapa eins og galagóa og pottóa í Afríku, lemúra á Madagaskar og lóríur í Suðaustur-Asíu. Haplorhini, eða þurrnefsprímatar, innihalda tarsíera (Mynd 29.42) og eiginlega apa, það er apa Nýja heimsins, apa Gamla heimsins, mannapa og menn. Almennt eru Strepsirrhini oftar næturdýr, hafa stærri lyktarskynsstöðvar í heilanum og eru smærri og með minni heila en anthropoid-prímatar. Haplorhini eru, með fáum undantekningum, dagdýr og treysta meira á sjón. Annar áhugaverður munur er að Strepsirrhini hafa ensímin sem þarf til að framleiða C-vítamín, en Haplorhini verða að fá C-vítamín úr fæðunni.

Þróun prímata
Fyrstu prímatalíku spendýrin eru kölluð frumprímatar. Þau voru svipuð íkornum og trjásnjáldrum að stærð og útliti. Fyrirliggjandi steingervingar, aðallega frá Norður-Afríku, eru mjög brotakenndir og frumprímatar eru því enn að mestu dularfullur hópur þar til fleiri steingervingar finnast. Þótt erfðafræðileg gögn bendi til þess að prímatar hafi greinst frá öðrum spendýrum fyrir um 85 milljónum ára, eru elstu þekktu prímatalíku spendýrin með tiltölulega trausta steingervingasögu frá því fyrir um 65 milljónum ára. Steingervingar á borð við frumprímatann Plesiadapis, þó ekki séu allir rannsakendur sammála um að hann hafi verið frumprímati, höfðu sum einkenni tanna og beinagrindar sameiginleg með sönnum prímötum. Þeir hafa fundist í Norður-Ameríku og Evrópu á nýlífsöld (Cenozoic) og dóu út undir lok eósentímabilsins, fyrir um 56 til 34 milljónum ára.
Fyrstu sönnu prímatarnir komu fram fyrir um 55 milljónum ára á eósentímabilinu. Þeir fundust í Norður-Ameríku, Evrópu, Asíu og Afríku og líktust núlifandi hálföpum eins og lemúrum. Þróunarbreytingar héldu áfram hjá þessum fyrstu prímötum; stærri heilar og augu og minni trýni voru ríkjandi þróun. Undir lok eósentímabilsins dóu margar fyrstu tegundir hálfapa út, annaðhvort vegna kólnandi loftslags eða samkeppni frá fyrstu öpunum.
Eiginlegir apar (anthropoids) þróuðust frá hálföpum á eósentímabilinu. Fyrir 40 milljónum ára benda gögn til þess að apar hafi verið til staðar bæði í Nýja heiminum, það er Suður-Ameríku, og Gamla heiminum, Afríku og Asíu. Apar Nýja heimsins eru einnig kallaðir Platyrrhini, sem vísar til breiðra nasa þeirra (Mynd 29.43). Apar Gamla heimsins eru kallaðir Catarrhini, sem vísar til mjórra nasa sem vísa niður. Enn ríkir talsverð óvissa um uppruna apa Nýja heimsins. Þegar Platyrrhini komu fram höfðu meginlönd Suður-Ameríku og Afríku rekið í sundur. Því er talið að apar hafi komið fram í Gamla heiminum og komist til Nýja heimsins annaðhvort með því að reka á gróðurflekunum eða fara yfir landbrýr. Vegna þessarar æxlunarlegu einangrunar gengu apar Nýja heimsins og apar Gamla heimsins í gegnum aðskilda aðlögunarkvíslun yfir milljónir ára. Apar Nýja heimsins eru allir trjábúar, en apar Gamla heimsins innihalda bæði trjábúa og tegundir sem lifa á jörðu niðri. Trjávenjur apa Nýja heimsins endurspeglast í því að flestar tegundir hafa griphala. Halar apa Gamla heimsins eru aldrei griphalar og eru oft smáir, og sumar tegundir hafa setþófa, það er þykknaða húðbletti á rassinum.

Mannapar þróuðust frá Catarrhini-öpum í Afríku um miðja nýlífsöld, fyrir um 25 milljónum ára. Mannapar eru almennt stærri en apar og hafa ekki hala. Allir mannapar geta hreyft sig í trjám, þótt margar tegundir verji mestum tíma sínum á jörðu niðri. Þegar apar ganga á fjórum fótum ganga þeir á lófunum, en mannapar styðja efri hluta líkamans á hnúunum. Mannapar eru greindari en apar og hafa stærri heila miðað við líkamsstærð. Mannöpum er skipt í tvo hópa. Minni mannapar tilheyra ættinni Hylobatidae, sem inniheldur gibbon-apa og síamanga. Stórir mannapar innihalda ættkvíslirnar Pan (simpansar og bonobo-apar), Gorilla (górillur), Pongo (órangútanar) og Homo (menn) (Mynd 29.44).

Gibbon-apar, sem eru mjög trjábundnir, gibbon-apar eru minni en stórir mannapar; þeir hafa lítinn kynjamun (það er að segja, kynin eru ekki áberandi ólík að stærð), þó að hjá sumum tegundum séu kynin ólík á litinn; og þeir hafa hlutfallslega lengri handleggi sem notaðir eru til að sveifla sér milli trjáa (Mynd 29.45 a). Tvær tegundir órangútana eru upprunnar á mismunandi eyjum í Indónesíu: Borneó (P. pygmaeus) og Súmötru (P. abelii). Þriðja órangútan-tegundin, Pongo tapanuliensis, var tilkynnt árið 2017 frá Batang Toru skóginum á Súmötru. Órangútanar eru trjábúar og lifa einir. Karlkyns dýr eru mun stærri en kvenkyns og hafa kinn- og hálspoka þegar þeir eru fullþroska. Górillur lifa allar í Mið-Afríku. Austur- og vesturstofnarnir eru viðurkenndir sem aðskildar tegundir, G. beringei og G. gorilla. Górillur sýna sterkan kynjamun, þar sem karldýr eru um það bil tvöfalt stærri en kvendýr. Hjá eldri karldýrum, kölluðum silfurbökum, verður hárið á bakinu hvítt eða grátt. SimPansar (Mynd 29.45 b) eru sú tegund sem talin er skyldust mönnum. Hins vegar er sú tegund sem er skyldust simPansanum bonobo-apinn. Erfðafræðileg gögn benda til þess að ættarlínur simpansa og manna hafi aðskist fyrir 5 til 7 milljónum ára, en ættarlínur simpansa (Pan troglodytes) og bonobo-apa (Pan paniscus) hafi aðskist fyrir um 2 milljónum ára. Simpansar og bonobo-apar lifa báðir í Mið-Afríku, en tegundirnar tvær eru aðskildar af Kongófljóti, sem er veruleg landfræðileg hindrun. Bonobo-apar eru grannvaxnari en simpansar, en hafa lengri fætur og meira hár á höfðinu. Hjá simpönsum auðkenna hvítir haladúskar unga, en bonobo-apar halda hvítum haladúskum sínum alla ævi. Bonobo-apar hafa einnig skærari raddir en simpansar. Simpansar eru árásargjarnari og drepa stundum dýr úr öðrum hópum, en bonobo-apar eru ekki þekktir fyrir að gera slíkt. Bæði simpansar og bonobo-apar eru alætur. Fæða órangútana og górilla inniheldur einnig fæðu úr mörgum uppsprettum, þó að ríkjandi fæðutegundir séu ávextir fyrir órangútana og lauf fyrir górillur.

Þróun mannsins
Ættin Hominidae innan ættbálks prímata inniheldur stóru mannapa og menn (Mynd 29.46). Vísbendingar úr steingervingasögunni og samanburður á DNA manna og simpansa benda til þess að menn og simpansar hafi greinst frá sameiginlegum forföður mannapa fyrir um sex milljónum ára. Nokkrar tegundir þróuðust út frá þeirri grein sem inniheldur menn, þótt okkar tegund sé sú eina sem lifir enn. Hugtakið hominíni er notað um þær tegundir sem þróuðust eftir þennan aðskilnað prímatalínunnar og vísar þannig til tegunda sem eru skyldari mönnum en simpönsum. Nokkur einkenni aðgreina menn frá öðrum mannöpum, þar á meðal tvífætni eða upprétt staða, aukin heilastærð og fullkomlega gagnstæður þumall sem getur snert litla fingur. Tvífættir hominínar innihalda nokkra hópa sem voru líklega hluti af þróunarlínu nútímamannsins, svo sem Australopithecus, Homo habilis og Homo erectus, og nokkra hópa sem eru ekki forfeður en má líta á sem „frændur“ nútímamanna, til dæmis neanderdalsmenn og Denisova-menn.
Það er erfitt að ákvarða hinar sönnu ættarlínur hominína. Áður fyrr, þegar tiltölulega fáir steingervingar hominína höfðu fundist, töldu sumir vísindamenn að með því að skoða þá í tímaröð, frá elstu til yngstu, mætti sýna fram á þróunarferlið frá fyrstu hominínum til nútímamanna. Á síðustu árum hafa hins vegar fundist margir nýir steingervingar og ljóst er að oft var meira en ein tegund á lífi á hverjum tíma og að margir af fundnum steingervingum (og nefndum tegundum) tákna tegundir hominína sem dóu út og eru ekki forfeður nútímamanna.
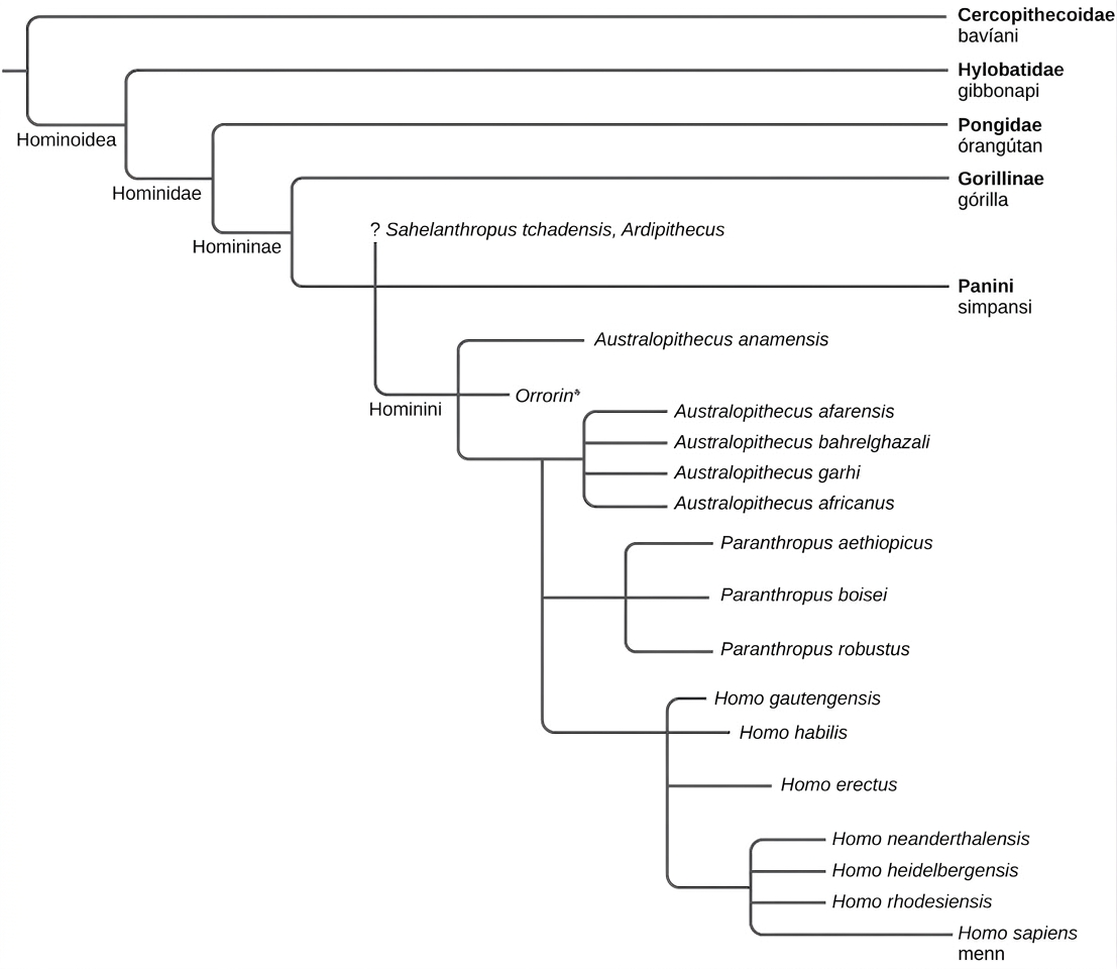
Mjög snemmbúnir hominínar
Þrjár tegundir mjög snemmbúinna hominína hafa verið í fréttum seint á 20. öld og snemma á þeirri 21.: Ardipithecus, Sahelanthropus og Orrorin. Yngsta tegundin af þessum þremur, Ardipithecus, fannst á tíunda áratugnum og er talin vera um 4,4 milljónum ára gömul. Þótt óvissa hafi verið um tvífætni fyrstu sýnanna, fundust fleiri sýni af Ardipithecus á árunum þar á eftir sem sýndu fram á að lífveran var tvífætt. Tvær mismunandi tegundir af Ardipithecus hafa verið greindar, A. ramidus og A. kadabba, en sýni þeirrar síðarnefndu eru eldri og talin vera 5,6 milljónum ára gömul. Staða þessarar ættkvíslar sem forföður manna er hins vegar óviss.
Sú elsta af þessum þremur, Sahelanthropus tchadensis, fannst árin 2001-2002 og hefur verið tímasett til fyrir tæplega sjö milljónum ára. Til er eitt sýni af þessari ættkvísl, höfuðkúpa sem fannst á yfirborði í Tsjad. Steingervingurinn, sem óformlega er kallaður „Toumai“, er blanda af frumstæðum og þróuðum einkennum, og óljóst er hvernig þessi steingervingur passar við þá mynd sem sameindagögn gefa, nefnilega að línan sem liggur til nútímamanna og nútímasimpansa hafi greinst í sundur fyrir um sex milljónum ára. Ekki er talið á þessari stundu að þessi tegund hafi verið forfaðir nútímamanna.
Yngri tegund, Orrorin tugenensis, sem er um sex milljóna ára gömul, er einnig tiltölulega nýleg uppgötvun og fannst árið 2000. Til eru nokkur sýni af Orrorin. Sum einkenni Orrorin eru líkari einkennum nútímamanna en einkennum suðurapa, þótt Orrorin sé mun eldri. Ef Orrorin er forfaðir manna gætu suðuraparnir ekki verið í beinni ættarlínu manna. Fleiri sýni af þessum tegundum gætu hjálpað til við að skýra hlutverk þeirra.
Snemmbúnir hominínar: Ættkvíslin Australopithecus
Australopithecus („suðurapi“) er ættkvísl hominína sem þróaðist í Austur-Afríku fyrir um fjórum milljónum ára og dó út fyrir um tveimur milljónum ára. Þessi ættkvísl vekur sérstakan áhuga þar sem talið er að okkar ættkvísl, Homo, hafi þróast frá sameiginlegum forföður með Australopithecus fyrir um tveimur milljónum ára, líklega eftir einhver millistig. Australopithecus hafði nokkur einkenni sem voru líkari einkennum stórra mannapa en nútímamanna. Til dæmis var kynjamunur meiri en hjá nútímamönnum. Karldýr voru allt að 50 prósent stærri en kvendýr, svipað og sést hjá nútímagórillum og órangútönum. Aftur á móti eru karlar nútímamanna um það bil 15 til 20 prósent stærri en konur. Heilastærð Australopithecus miðað við líkamsmassa var einnig minni en hjá nútímamönnum og líkari því sem sést hjá stórum mannöpum. Lykileinkenni sem Australopithecus átti sameiginlegt með nútímamönnum var tvífætni, þótt líklegt sé að Australopithecus hafi einnig varið tíma í trjám. Fótspor hominína, svipuð sporum nútímamanna, fundust í Laetoli í Tansaníu og voru tímasett til 3,6 milljóna ára. Þau sýndu að hominínar á tímum Australopithecus gengu uppréttir.
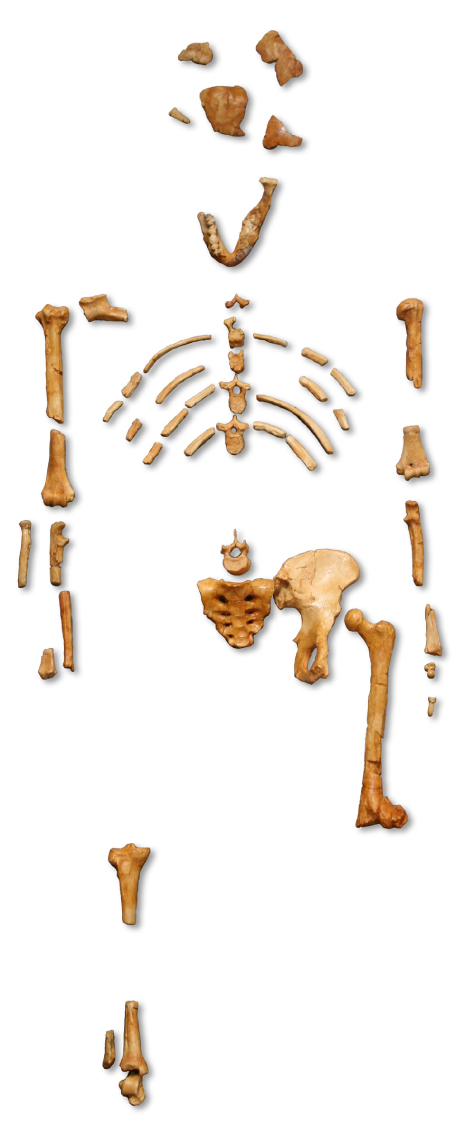
Til voru nokkrar tegundir af Australopithecus, sem oft eru kallaðar suðurapar. Australopithecus anamensis lifði fyrir um 4,2 milljónum ára. Meira er vitað um aðra snemmbúna tegund, Australopithecus afarensis, sem lifði fyrir 3,9 til 2,9 milljónum ára. Þessi tegund sýnir þróunartilhneigingu í þróun mannsins: minnkun tanna og kjálka. A. afarensis (Mynd 29.47a) hafði minni vígtennur og jaxla en apar, en tennurnar voru stærri en hjá nútímamönnum. Heilastærð hennar var 380 til 450 rúmsentimetrar, um það bil á stærð við heila nútímasimpansa. Hún hafði einnig framstæðan kjálka, það er hlutfallslega lengri kjálka en nútímamenn. Um miðjan áttunda áratuginn fannst steingervingur af fullorðnu kvendýri af tegundinni A. afarensis í Afar-héraði í Eþíópíu og var hann tímasettur til 3,24 milljóna ára (Mynd 29.48). Steingervingurinn, sem óformlega er kallaður „Lucy“, er mikilvægur vegna þess að hann var heillegasti steingervingur suðurapa sem fundist hafði, en um 40 prósent beinagrindarinnar fundust.

Australopithecus africanus lifði fyrir tveimur til þremur milljónum ára. Tegundin var grannvaxin og tvífætt en hafði sterkbyggð handleggsbein og gæti, líkt og aðrir snemmbúnir hominínar, hafa varið verulegum tíma í trjám. Heili hennar var stærri en heili A. afarensis, eða um 500 rúmsentimetrar, sem er aðeins minna en þriðjungur af stærð heila nútímamanns. Tvær aðrar tegundir, Australopithecus bahrelghazali og Australopithecus garhi, hafa bæst í hóp suðurapa á undanförnum árum. A. bahrelghazali er óvenjuleg þar sem hún er eini suðurapinn sem fundist hefur í Mið-Afríku.
Blindgata: Ættkvíslin Paranthropus
Suðuraparnir voru tiltölulega grannvaxnir og með tennur sem hentuðu fyrir mjúka fæðu. Á síðustu árum hafa fundist steingervingar hominína með annars konar líkamsbyggingu sem tímasettir eru til fyrir um 2,5 milljónum ára. Þessir hominínar, af ættkvíslinni Paranthropus, voru vöðvastæltir, 1,3 til 1,4 metrar á hæð og með stórar jaxlatennur. Jaxlar þeirra sýndu mikið slit, sem bendir til þess að þeir hafi lifað á grófri og trefjaríkri jurtafæðu, ólíkt suðuröpunum sem voru að hluta til kjötætur. Paranthropus inniheldur Paranthropus robustus frá Suður-Afríku, og Paranthropus aethiopicus og Paranthropus boisei frá Austur-Afríku. Hominíðarnir í þessari ættkvísl dóu út fyrir meira en einni milljón ára og eru ekki taldir vera forfeður nútímamanna, heldur meðlimir þróunargreinar á ættartré hominína sem létu enga afkomendur eftir sig.
Snemmbúnir hominínar: Ættkvíslin Homo
Mannsættkvíslin Homo kom fyrst fram fyrir 2,5 til 3 milljónum ára. Í mörg ár voru steingervingar tegundar sem kallast H. habilis elstu dæmin um ættkvíslina Homo, en árið 2010 fannst ný tegund sem kallast Homo gautengensis og gæti verið eldri. Samanborið við A. africanus hafði H. habilis nokkur einkenni sem voru líkari nútímamönnum. H. habilis hafði kjálka sem var minna framstæður en hjá suðuröpum og stærri heila, 600 til 750 rúmsentimetra. Hins vegar hélt H. habilis sumum einkennum eldri hominína, svo sem löngum handleggjum. Nafnið H. habilis merkir „hinn handlagni maður“ og vísar til steinverkfæra sem fundist hafa með leifum hans.
Tengill í námsefni
Tengill í námsefni: Horfðu á þetta myndband um steingervingafræðinginn Briana Pobiner frá Smithsonian þar sem hún útskýrir tengslin milli kjötáts hominína og þróunar mannsins.
H. erectus kom fram fyrir um 1,8 milljónum ára (Mynd 29.49). Talið er að tegundin hafi átt uppruna sinn í Austur-Afríku og hún var fyrsti hominíninn sem fluttist út úr Afríku. Steingervingar af H. erectus hafa fundist á Indlandi, í Kína, á Jövu og í Evrópu og voru áður þekktir sem „Jövumaðurinn“ eða „Pekingmaðurinn“. H. erectus hafði mörg einkenni sem voru líkari nútímamönnum en einkennum H. habilis. H. erectus var stærri en eldri hominínar, náði allt að 1,85 metra hæð og vó allt að 65 kílógrömm, sem er svipað og hjá nútímamönnum. Kynjamunur var minni en hjá eldri tegundum; karldýr voru 20 til 30 prósent stærri en kvendýr, sem er nálægt stærðarmuninum hjá okkar eigin tegund. H. erectus hafði stærri heila en eldri tegundir, 775 til 1.100 rúmsentimetra, samanborið við 1.130 til 1.260 rúmsentimetra hjá nútímamönnum. H. erectus hafði einnig nef með niðurvísandi nösum líkt og nútímamenn, fremur en framvísandi nasir eins og finnast hjá öðrum prímötum. Lengri, niðurvísandi nasir gera kleift að hita kalt loft áður en það fer í lungun og gætu hafa verið aðlögun að kaldara loftslagi. Gripir sem fundist hafa með steingervingum H. erectus benda til þess að þetta hafi verið fyrsti hominíninn sem notaði eld, veiddi og hafði bækistöðvar. Almennt er talið að H. erectus hafi lifað þar til fyrir um 50.000 árum.

Menn: Homo sapiens
Nokkrar tegundir, stundum kallaðar fornlegir Homo sapiens, þróuðust greinilega frá H. erectus og hófst það ferli fyrir um 500.000 árum. Þessar tegundir eru meðal annars Homo heidelbergensis, Homo rhodesiensis og Homo neanderthalensis. Þessir fornlegu H. sapiens höfðu heilastærð svipaða og nútímamenn, að meðaltali 1.200 til 1.400 rúmsentimetrar. Þeir voru frábrugðnir nútímamönnum að því leyti að þeir höfðu þykka höfuðkúpu, áberandi augabrúnakamb og höku sem vísaði inn á við. Sumar þessara tegunda lifðu þar til fyrir 30.000 til 10.000 árum og voru samtíða nútímamönnum (Mynd 29.50).

Talsverðar deilur eru um uppruna líffærafræðilega nútímalegra manna, Homo sapiens sapiens. Eins og rætt var um fyrr fluttist H. erectus út úr Afríku og inn í Asíu og Evrópu í fyrstu stóru fólksflutningabylgjunni fyrir um 1,5 milljónum ára. Talið er að nútímamenn hafi komið fram í Afríku af H. erectus og flust út úr Afríku fyrir um 100.000 árum í annarri stórri fólksflutningabylgju. Síðan hafi nútímamenn komið í stað tegunda H. erectus sem höfðu flust til Asíu og Evrópu í fyrstu bylgjunni.
Þessi þróunartímalína er studd af sameindafræðilegum sönnunargögnum. Ein nálgun við að rannsaka uppruna nútímamanna er að skoða hvatbera-DNA (mtDNA) frá stofnum um allan heim. Þar sem fóstur þroskast úr eggi sem inniheldur hvatbera móðurinnar, sem hafa sitt eigið DNA utan kjarna, erfist mtDNA alfarið í kvenlegg. Nú er hægt að nota stökkbreytingar í mtDNA til að áætla tímalínu erfðafræðilegs aðskilnaðar. Niðurstöðurnar benda til þess að allir nútímamenn hafi mtDNA sem er erft frá sameiginlegum forföður sem lifði í Afríku fyrir um 160.000 árum. Önnur nálgun á sameindafræðilegum skilningi á þróun mannsins er að skoða Y-litninginn, sem erfist frá föður til sonar. Þessi gögn benda til þess að allir núlifandi karlar hafi erft Y-litning frá karlmanni sem lifði í Afríku fyrir um 140.000 árum.
Rannsóknir á hvatbera-DNA leiddu til greiningar á annarri manntegund eða undirtegund, Denisova-mönnum. DNA úr tönnum og fingurbeinum bentu til tveggja atriða. Í fyrsta lagi var hvatbera-DNA frábrugðið því sem finnst bæði hjá nútímamönnum og neanderdalsmönnum. Í öðru lagi benti erfðamengis-DNA til þess að Denisova-menn deildu sameiginlegum forföður með neanderdalsmönnum. Gen frá bæði neanderdalsmönnum og Denisova-mönnum hafa fundist í stofnum nútímamanna, sem bendir til þess að blöndun hafi átt sér stað milli hópanna þriggja á hluta útbreiðslusvæðis þeirra.