7.7 Stjórnun frumuöndunar
7.7 Stjórnun frumuöndunar
Námsmarkmið
Í lok þessa kafla muntu geta gert eftirfarandi:
- Lýst hvernig afturvirk hömlun hefur áhrif á myndun milliefnis eða myndefnis í efnaskiptaferli
- Borið kennsl á þann búnað sem stjórnar hraða rafeindaflutnings um rafeindaflutningskeðjuna
Frumuöndun þarf að vera stjórnað svo fruman fái hæfilegt magn orku í formi ATP. Fruman þarf einnig að mynda ýmis milliefni sem eru notuð í uppbyggingu og niðurbroti stórsameinda. Án stjórnunar myndu efnaskiptahvörf fljótt stöðvast þegar fram- og afturhvörf næðu jafnvægi. Þá yrðu auðlindir frumunnar nýttar á óhentugan hátt. Fruma þarf ekki alltaf að mynda eins mikið ATP og hún getur; stundum þarf hún að beina sumum milliefnum inn í ferla sem mynda amínósýrur, prótein, glýkógen, lípíð og kjarnsýrur. Í stuttu máli þarf fruman að stjórna efnaskiptum sínum.
Stjórnunarferlar
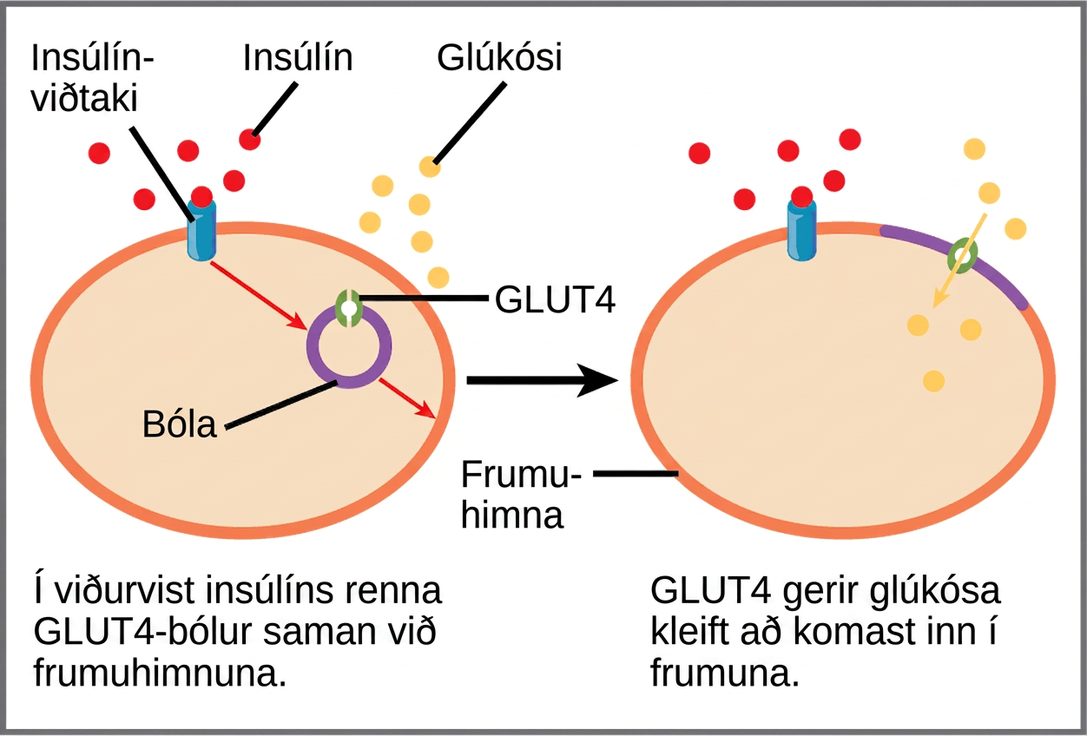
Ýmsir búnaðurar eru notaðir til að stjórna frumuöndun. Einhvers konar stjórnun er til staðar á hverju stigi glúkósaefnaskipta. Aðgangi glúkósa að frumunni má stjórna með GLUT-próteinum (glúkósaflutningspróteinum), sem flytja glúkósa (mynd 7.20). Mismunandi gerðir GLUT-próteina stjórna flutningi glúkósa inn í frumur tiltekinna vefja.
Sumum hvörfum er stjórnað með því að hafa tvö mismunandi ensím, eitt fyrir hvora stefnu afturkræfs hvarfs. Hvörf sem aðeins eitt ensím hvatar geta náð jafnvægi og þá stöðvast hvarfið. Ef tvö mismunandi ensím, hvort sértækt fyrir sína stefnu, eru hins vegar nauðsynleg fyrir afturkræft hvarf eykst möguleikinn á að stjórna hraða hvarfsins og jafnvægi næst ekki.
Fjölda ensíma í hverjum efnaskiptaferli, sérstaklega ensíminu sem hvetur fyrsta skuldbindandi hvarf ferilsins, er stjórnað með því að sameind binst fjarvirku setri á próteininu. Sameindirnar sem oftast gegna þessu hlutverki eru kirnin ATP, ADP, AMP, NAD+ og NADH. Þessir stjórnarþættir, fjarvirkir áhrifavaldar, geta aukið eða minnkað virkni ensíms eftir aðstæðum. Fjarvirkur áhrifavaldur breytir þrívíddarbyggingu ensímsins og hefur venjulega áhrif á lögun hvarfstöðvarinnar. Þessi breyting á byggingu próteinsins, ensímsins, eykur eða minnkar sækni þess í hvarfmiðinn og hækkar eða lækkar þar með hraða hvarfsins. Bindingin sendir ensíminu boð og getur aukið eða minnkað virkni þess; þannig verður til afturvirkur stjórnbúnaður. Þessi tegund afturvirkrar stjórnunar virkar svo lengi sem efnið sem hefur áhrif á hana er bundið ensíminu. Þegar heildarstyrkur efnisins minnkar flæðir það frá próteininu og stjórnuninni er aflétt.
Stjórnun niðurbrotsferla
Ensím, prótein, rafeindaferjur og dælur sem gegna hlutverki í glýkólýsu, sítrónusýruhringnum og rafeindaflutningskeðjunni hafa tilhneigingu til að hvata óafturkræf hvörf. Með öðrum orðum: ef upphafshvarfið fer fram er ferillinn skuldbundinn til að halda áfram með þau hvörf sem eftir eru. Hvort tiltekin ensímvirkni er sett af stað fer eftir orkuþörf frumunnar, eins og hún endurspeglast í magni ATP, ADP og AMP.
Glýkólýsa
Stjórnun glýkólýsu hefst með fyrsta ensíminu í ferlinum, hexókínasa (mynd 7.21). Þetta ensím hvetur fosfórun glúkósa, sem undirbýr efnasambandið fyrir klofning í síðara skrefi. Neikvætt hlaðni fosfathópurinn í sameindinni kemur einnig í veg fyrir að sykurinn fari út úr frumunni. Þegar hexókínasi er hamlaður flæðir glúkósi út úr frumunni og verður ekki hvarfmið í öndunarferlum í þeim vef. Myndefni hexókínasahvarfsins er glúkósa-6-fosfat, sem safnast upp þegar síðara ensím, fosfófrúktókínasi, er hamlað.
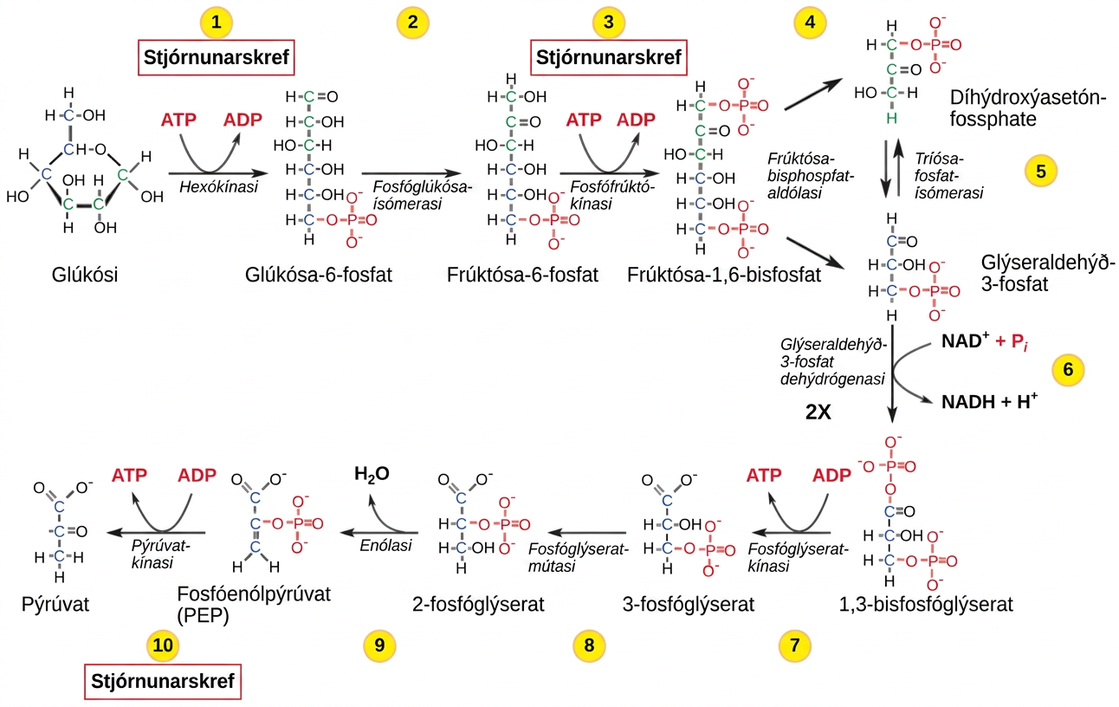
Fosfófrúktókínasi er helsta stjórnaða ensímið í glýkólýsu. Mikið magn ATP eða sítrats, eða lægra og súrara pH-gildi, minnkar virkni ensímsins. Styrkur sítrats getur aukist vegna hindrunar í sítrónusýruhringnum. Gerjun, sem myndar lífrænar sýrur eins og mjólkursýru, skýrir oft aukið sýrustig í frumu; myndefni gerjunar safnast þó yfirleitt ekki upp í frumum.
Síðasta skref glýkólýsu er hvatað af pýrúvatkínasa. Pýrúvatið sem myndast getur haldið áfram í niðurbrot eða verið breytt í amínósýruna alanín. Ef ekki er þörf á meiri orku og alanín er til í nægu magni er ensímið hamlað. Virkni ensímsins eykst þegar magn frúktósa-1,6-bisfosfats eykst. Mundu að frúktósa-1,6-bisfosfat er milliefni í fyrri hluta glýkólýsu. Stjórnun pýrúvatkínasa felur í sér fosfórun af völdum kínasa, sem leiðir til minna virks ensíms. Affosfórun með fosfatasa endurvirkjar það. Pýrúvatkínasa er einnig stjórnað af ATP, sem hefur neikvæð fjarvirk áhrif.
Ef þörf er á meiri orku breytist meira pýrúvat í asetýl-CoA fyrir tilstilli pýrúvatdehýdrógenasa. Ef annaðhvort asetýlhópar eða NADH safnast upp er minni þörf fyrir hvarfið og hraðinn minnkar. Pýrúvatdehýdrógenasa er einnig stjórnað með fosfórun: kínasi fosfórar ensímið og gerir það óvirkt, en fosfatasi endurvirkjar það. Kínasinn og fosfatasinn eru einnig undir stjórnun.
Sítrónusýruhringurinn
Sítrónusýruhringnum er stjórnað með ensímunum sem hvetja hvörfin sem mynda fyrstu tvær NADH-sameindirnar (mynd 7.11). Þessi ensím eru ísósítratdehýdrógenasi og α-ketóglútaratdehýdrógenasi. Þegar nægilegt magn ATP og NADH er til staðar minnkar hraði þessara hvarfa. Þegar þörf er á meira ATP, sem endurspeglast í hækkandi magni ADP, eykst hraðinn. α-ketóglútaratdehýdrógenasi verður einnig fyrir áhrifum af magni súkkínýl-CoA, milliefnis sem kemur síðar í hringnum, og það veldur minni virkni. Minni hraði ferilsins á þessum stað er ekki endilega neikvæður, því aukið magn α-ketóglútarats sem sítrónusýruhringurinn notar ekki getur nýst frumunni við myndun amínósýrunnar glútamats.
Rafeindaflutningskeðjan
Sértæk ensím rafeindaflutningskeðjunnar verða ekki fyrir áhrifum af afturvirkri hömlun, en magn ADP og ATP hefur áhrif á hraða rafeindaflutnings um keðjuna. Aukin ATP-notkun í frumu kemur fram sem uppsöfnun ADP. Þegar ATP-notkun minnkar lækkar styrkur ADP og ATP byrjar að safnast upp í frumunni. Þessi breyting á hlutfallslegum styrk ADP miðað við ATP veldur því að fruman hægir á rafeindaflutningskeðjunni.
Sjá samantekt á afturvirkri stjórnun frumuöndunar í töflu 7.1.
| Ferill | Ensím sem verður fyrir áhrifum | Hækkað magn áhrifavalds | Áhrif á virkni ferils |
|---|---|---|---|
| glýkólýsa | hexókínasi | glúkósa-6-fosfat | minnkun |
| fosfófrúktókínasi | lítil orkuhleðsla (ATP, AMP), frúktósa-6-fosfat fyrir tilstilli frúktósa-2,6-bisfosfats | aukning | |
| mikil orkuhleðsla (ATP, AMP), sítrat, súrt pH-gildi | minnkun | ||
| pýrúvatkínasi | frúktósa-1,6-bisfosfat | aukning | |
| mikil orkuhleðsla (ATP, AMP), alanín | minnkun | ||
| umbreyting pýrúvats í asetýl-CoA | pýrúvatdehýdrógenasi | ADP, pýrúvat | aukning |
| asetýl-CoA, ATP, NADH | minnkun | ||
| sítrónusýruhringur | ísósítratdehýdrógenasi | aukning | |
| minnkun | |||
| α-ketóglútaratdehýdrógenasi | kalsíumjónir, ADP | aukning | |
| ATP, NADH, súkkínýl-CoA | minnkun | ||
| rafeindaflutningskeðja | aukning | ||
| minnkun |