7.4 Oxunarfosfórun
7.4 Oxunarfosfórun
Námsmarkmið
Í lok þessa kafla muntu geta gert eftirfarandi:
- Lýst hvernig rafeindir fara í gegnum rafeindaflutningskeðjuna og útskýrt hvað verður um orkustig þeirra í þessu ferli
- Útskýrt hvernig róteindastigull (H⁺) myndast og hvernig rafeindaflutningskeðjan viðheldur honum
Þú hefur nýlega lesið um tvo ferla í niðurbroti glúkósa, glýkólýsu og sítrónusýruhringinn, sem mynda ATP. Mest af ATP sem myndast við loftháð niðurbrot glúkósa myndast þó ekki beint í þessum ferlum. Þess í stað verður það til í ferli sem hefst þegar rafeindir flytjast eftir röð rafeindaferja sem gangast undir oxunar-afoxunarhvörf. Ferlið veldur því að vetnisjónir safnast upp í millihimnurýminu. Þannig myndast styrkstigull þar sem vetnisjónir flæða úr millihimnurýminu inn í hvatberamerginn í gegnum ATP-syntasa. Straumur vetnisjóna knýr hvatavirkni ATP-syntasa, sem fosfórar ADP og myndar ATP.
Rafeindaflutningskeðja
Rafeindaflutningskeðjan (mynd 7.12) er síðasti hluti loftháðrar öndunar og er eini hluti glúkósaefnaskipta sem notar súrefni úr andrúmslofti. Súrefni flæðir stöðugt inn í plöntuvefi, yfirleitt um loftaugu, og einnig inn í sveppi og bakteríur; hjá dýrum berst súrefni hins vegar inn í líkamann um ýmis öndunarkerfi. Rafeindaflutningur er röð oxunar-afoxunarhvarfa sem líkist boðhlaupi eða fötukeðju: rafeindir berast hratt frá einum hluta til hins næsta, allt að enda keðjunnar þar sem þær afoxa sameindasúrefni og mynda vatn ásamt tengdum róteindum. Fjórir próteinflókar, merktir I til IV á mynd 7.12, ásamt hreyfanlegum aukarafeindaferjum mynda rafeindaflutningskeðjuna. Rafeindaflutningskeðjan er í mörgum eintökum í innri hvatberahimnu heilkjörnunga og í frumuhimnu dreifkjörnunga.
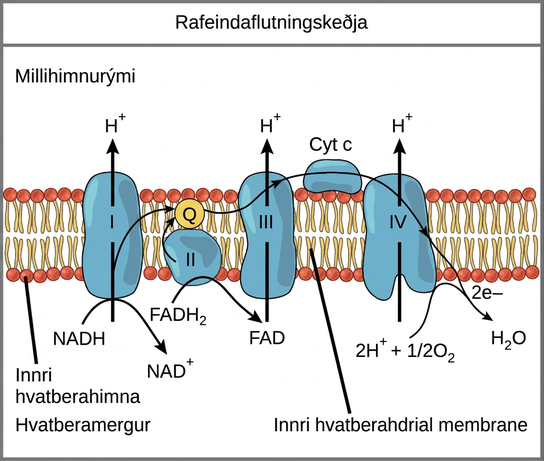
Flóki I
Fyrst berast tvær rafeindir til fyrsta flókans með NADH. Þessi flóki, merktur I, er samsettur úr flavínmónónúkleótíði (FMN) og próteini sem inniheldur járn-brennisteinsklasa (Fe-S). FMN, sem er afleitt af B2-vítamíni, einnig kallað ríbóflavín, er einn af nokkrum hjálparhópum eða hjálparþáttum í rafeindaflutningskeðjunni. Hjálparhópur er sameind sem er ekki prótein en er nauðsynleg fyrir virkni próteins. Hjálparhópar eru lífrænar eða ólífrænar sameindir, ekki peptíð, sem bindast próteinum og auðvelda virkni þeirra. Til hjálparhópa teljast kóensím, sem eru hjálparhópar ensíma. Ensímið í flóka I er NADH-dehýdrógenasi og er samsett úr 44 aðskildum fjölpeptíðkeðjum. Flóki I getur dælt fjórum vetnisjónum yfir himnuna frá mergnum inn í millihimnurýmið; þannig myndast vetnisjónastigullinn milli hólfanna tveggja sem innri hvatberahimnan aðskilur og honum er viðhaldið.
Q og flóki II
Flóki II tekur beint við FADH2, sem fer ekki í gegnum flóka I. Efnasambandið sem tengir fyrsta og annan flóka við þann þriðja er ubíkínón (hjálparensím Q). Q-sameindin er fituleysanleg og hreyfist frjálst um vatnsfælinn kjarna himnunnar. Þegar hún hefur afoxast (QH2) skilar ubíkínón rafeindum sínum til næsta flóka í rafeindaflutningskeðjunni. Q tekur við rafeindum frá NADH úr flóka I og rafeindum frá FADH2 úr flóka II. Þetta ensím og FADH2 mynda lítinn flóka sem skilar rafeindum beint til rafeindaflutningskeðjunnar og fer fram hjá fyrsta flókanum. Þar sem þessar rafeindir fara fram hjá fyrsta flókanum og knýja því ekki róteindadæluna í honum myndast færri ATP-sameindir úr rafeindum FADH2. Fjöldi ATP-sameinda sem fæst að lokum er í beinu hlutfalli við fjölda róteinda sem dælt er yfir innri hvatberahimnuna.
Flóki III
Þriðji flókinn er samsettur úr cýtókrómi b, öðru Fe-S-próteini, Rieske-setri (2Fe-2S-setri) og cýtókróm c-próteinum. Þessi flóki er einnig kallaður cýtókróm-oxídóredúktasi. Cýtókrómprótein hafa hem sem hjálparhóp. Hemsameindin líkist heminu í blóðrauða en ber rafeindir, ekki súrefni. Þess vegna afoxast og oxast járnjónin í kjarna hennar þegar hún flytur rafeindirnar áfram og sveiflast milli oxunarstiga: Fe2+ (afoxað) og Fe3+ (oxað). Hemsameindir í cýtókrómum hafa lítillega mismunandi eiginleika vegna áhrifa próteinanna sem bindast þeim og það gefur hverjum flóka aðeins ólíka eiginleika. Flóki III dælir róteindum yfir himnuna og sendir rafeindir sínar til cýtókróms c, sem flytur þær til fjórða flókans af próteinum og ensímum. Cýtókróm c tekur við rafeindum frá Q; en á meðan Q ber rafeindapör getur cýtókróm c aðeins tekið við einni rafeind í einu.
Flóki IV
Fjórði flókinn er samsettur úr cýtókrómpróteinum c, a og a3. Þessi flóki inniheldur tvo hemhópa, einn í hvoru cýtókróminu a og a3, og þrjár koparjónir, par af Cuₐ og eina Cuᵦ í cýtókrómi a3. Cýtókrómin halda súrefnissameind mjög þétt milli járn- og koparjónanna þar til súrefnið er fullkomlega afoxað með því að taka við tveimur rafeindum. Afoxaða súrefnið tekur síðan upp tvær vetnisjónir úr umhverfinu og myndar vatn (H2O). Fjarlæging vetnisjónanna úr kerfinu stuðlar að jónastiglinum sem er grunnur efnaosmósu.
Efnaosmósa
Í efnaosmósu er frjáls orka úr röð oxunar-afoxunarhvarfa sem lýst var hér að framan notuð til að dæla vetnisjónum (róteindum) yfir hvatberahimnuna. Ójöfn dreifing H+-jóna yfir himnuna myndar bæði styrkstigul og rafstigul, og þar með rafefnastigul, vegna jákvæðrar hleðslu vetnisjónanna og uppsöfnunar þeirra öðrum megin himnunnar.
Ef himnan væri stöðugt opin fyrir einföldu flæði vetnisjóna myndu jónirnar leita aftur yfir í merginn, knúnar áfram af styrkmuninum sem myndar rafefnastigul þeirra. Rifjið upp að margar jónir geta ekki flætt í gegnum óskautuð svæði fosfólípíðhimna án hjálpar jónaganga. Á sama hátt geta vetnisjónir í mergrýminu aðeins farið í gegnum innri hvatberahimnuna með hjálp íhimnupróteins sem kallast ATP-syntasi (mynd 7.13). Þetta flókna prótein virkar eins og lítill rafall sem snýst fyrir tilstilli krafts vetnisjónanna þegar þær flæða í gegnum það niður rafefnastigul sinn. Snúningur hluta þessarar sameindavélar auðveldar viðbót fosfats við ADP og myndar ATP með því að nýta stöðuorku vetnisjónastigulsins.
Efnaosmósa (mynd 7.14) er notuð til að mynda 90 prósent af ATP sem verður til við loftháð niðurbrot glúkósa. Hún er einnig aðferðin sem notuð er í ljóshvörfum ljóstillífunar til að beisla orku sólarljóss í ljósfosfórun. Rifjið upp að ATP-myndun með efnaosmósu í hvatberum kallast oxunarfosfórun. Heildarniðurstaða þessara hvarfa er myndun ATP úr orku rafeindanna sem voru fjarlægðar af vetnisatómum. Þessi atóm voru upphaflega hluti af glúkósasameind. Í lok ferilsins eru rafeindirnar notaðar til að afoxa súrefnissameind í súrefnisjónir. Aukarafeindirnar á súrefninu draga til sín vetnisjónir (róteindir) úr umhverfinu og vatn myndast. Þannig er súrefni lokarafeindaþegi í rafeindaflutningskeðjunni.
Afrakstur ATP
Fjöldi ATP-sameinda sem myndast við niðurbrot glúkósa er breytilegur. Til dæmis er misjafnt milli tegunda hversu mörgum vetnisjónum flókar rafeindaflutningskeðjunnar geta dælt í gegnum himnuna. Önnur uppspretta breytileika er ferjun rafeinda yfir himnur hvatberanna. NADH sem myndast við glýkólýsu kemst ekki auðveldlega inn í hvatbera. Því eru rafeindir teknar upp inni í hvatberum af annaðhvort NAD+ eða FAD+. Eins og þú hefur lært áður geta þessar FAD+-sameindir flutt færri jónir; þar af leiðandi myndast færri ATP-sameindir þegar FAD+ virkar sem ferja. NAD+ er notað sem rafeindaferja í lifur og FAD+ virkar í heila.
Annar þáttur sem hefur áhrif á afrakstur ATP-sameinda úr glúkósa er að milliefni í þessum ferlum eru einnig notuð í öðrum tilgangi. Niðurbrot glúkósa tengist ferlum sem byggja upp eða brjóta niður öll önnur lífefnafræðileg efnasambönd í frumum og niðurstaðan er nokkuð flóknari en þær kjöraðstæður sem lýst hefur verið hingað til. Til dæmis fara aðrar sykrur en glúkósi inn í glýkólýsuferilinn til orkuvinnslu. Auk þess eru fimm kolefna sykrurnar sem mynda kjarnsýrur gerðar úr milliefnum í glýkólýsu. Ákveðnar ónauðsynlegar amínósýrur er hægt að mynda úr milliefnum bæði glýkólýsu og sítrónusýruhringsins. Lípíð, svo sem kólesteról og þríglýseríð, eru einnig gerð úr milliefnum í þessum ferlum, og bæði amínósýrur og þríglýseríð eru brotin niður til orku með þessum ferlum. Í heild vinna þessir glúkósaniðurbrotsferlar í lifandi kerfum um 34 prósent af orkunni sem er í glúkósa en afgangurinn losnar sem varmi.