26.1 Þróun fræplantna
26.1 Þróun fræplantna
Námsmarkmið
Í lok þessa hluta munt þú geta gert eftirfarandi:
Fyrstu plönturnar sem námu land voru líklega skyldar forfeðrum nútímamosa, sem taldir eru hafa komið fram fyrir um 500 milljónum ára. Á eftir þeim komu lifrarmosar og frumstæðar æðplöntur, byrkningar, sem nútímaburknar eru komnir af. Lífsferill mosa og byrkninga einkennist af ættliðaskiptum, sem einnig sjást hjá berfrævingum og dulfrævingum. Það sem aðgreinir mosa og byrkninga frá berfrævingum og dulfrævingum er hins vegar þörf þeirra fyrir vatn við æxlun. Til að ljúka lífsferli mosa og byrkninga þarf vatn, því karlkynliðurinn losar svipubornar sáðfrumur sem verða að synda til kvenkynfrumunnar eða eggsins og frjóvga það. Eftir frjóvgun skiptir okfruman sér og vex í tvílitna grólið, sem myndar síðan gróhirslur. Í gróhirslunum fara móðurfrumur í meiósu og mynda einlitna gró. Þegar gró losna í hentugu umhverfi spíra þau og ný kynslóð kynliða myndast.
Hjá fræplöntum leiddi þróunin til ríkjandi gróliðskynslóðar og samsvarandi minnkunar á kynliðnum, úr áberandi byggingu í smásæjan frumuklasa sem er lokaður inni í vefjum gróliðsins. Þótt lægri æðplöntur, svo sem jafnar og burknar, séu að mestu jafngróa og myndi aðeins eina tegund gróa, eru allar fræplöntur misgróa og mynda tvær tegundir gróa: stórgró, sem eru kvenleg, og smágró, sem eru karlleg. Stórgró þroskast í kvenkynliði sem mynda egg, og smágró þroskast í karlkynliði sem mynda sáðfrumur. Þar sem þroski kynliðanna er háður vatni og næringu frá vefjum ríkjandi gróliðsins eru þeir ekki sjálfstætt lifandi eins og kynliðir frælausra æðplantna.
Forfeður misgróa frælausra plantna, sem nútímaplöntur á borð við jafnaættkvíslina Selaginella sýna dæmi um, eru taldir þróunarlegir fyrirrennarar fræplantna. Í lífsferli Selaginella þroskast bæði karl- og kvengróhirslur innan sama stöngullíka gróaxins. Í hverri karlgróhirslu myndast mörg smágró við meiósu. Hvert smágró myndar litla frjóhirslu inni í gróhylki. Þegar hún þroskast losnar hún úr gróaxinu og myndar margar svipubornar sáðfrumur sem fara síðan út úr gróhylkinu. Í kvengróhirslu fer ein stórgrómóðurfruma í meiósu og myndar fjögur stórgró. Kynliðir þroskast innan hvers stórgrós og samanstanda af vefjamassa sem nærir síðar fósturvísinn og nokkrum eggflöskum. Kvenkynliðurinn getur verið áfram inni í leifum gróveggjarins í stórgróhirslunni þar til frjóvgun hefur átt sér stað og fósturvísirinn byrjar að þroskast. Þessi samsetning fósturvísis og næringarfrumna er nokkuð frábrugðin byggingu fræs, því nærandi fræhvíta í fræi myndast úr einni frumu en ekki mörgum frumum.
Bæði fræ og frjókorn aðgreina fræplöntur frá frælausum æðplöntum. Þessar nýjungar gerðu fræplöntum kleift að draga úr eða losna við háðni sína á vatni við frjóvgun kynfrumna og þroska fósturvísis, og nema þurrlendi. Frjókorn eru karlkynliðir sem innihalda sáðfrumur, kynfrumur plöntunnar. Litlu einlitna (1n) frumurnar eru umluktar verndandi hjúp sem kemur í veg fyrir þornun og vélrænan skaða. Frjókorn geta borist langt frá upprunalegum grólið og dreift genum plöntunnar. Fræ veita fósturvísinum vernd, næringu og leið til að haldast í dvala í tugi eða jafnvel þúsundir ára, svo spírun geti átt sér stað þegar vaxtarskilyrði eru hagstæð. Þannig gera fræ plöntum kleift að dreifa næstu kynslóð bæði í rúmi og tíma. Með slíkum þróunarlegum kostum hafa fræplöntur orðið farsælasti og kunnuglegasti hópur plantna.
Báðar aðlaganirnar víkkuðu út landnám plantna sem hófst með mosum og forfeðrum þeirra. Steingervingar staðsetja elstu greinilegu fræplönturnar fyrir um 350 milljónum ára. Fyrstu áreiðanlegu heimildir um berfrævinga tímasetja tilkomu þeirra við mörk devon- og kolatímabilsins, fyrir um 319 milljónum ára (Mynd 26.2). Á undan berfrævingum komu frumberfrævingar, fyrstu nöktu fræplönturnar, sem komu fram fyrir um 380 til 390 milljónum ára. Frumberfrævingar voru millihópur plantna sem líktust barrtrjám í útliti, því þeir mynduðu við með síðvexti æðvefja, en þeir æxluðust enn eins og burknar og losuðu gró út í umhverfið. Að minnsta kosti sumar tegundir voru misgróa. Frumberfrævingar, eins og hinn útdauði Archaeopteris, sem ekki má rugla saman við fornfuglinn Archaeopteryx, voru ríkjandi í skógum síðdevontímabilsins. Við upphaf miðlífsaldar, á trías fyrir um 240 milljónum ára, og um miðbik hennar, á júra fyrir um 205 milljónum ára, voru sannir berfrævingar hins vegar orðnir ríkjandi í landslaginu. Dulfrævingar tóku fram úr berfrævingum um miðbik krítartímabilsins, fyrir um 100 milljónum ára, seint á miðlífsöld, og í dag eru þeir algengasti og líffræðilega fjölbreyttasti plöntuhópurinn í flestum lífbeltum á landi.
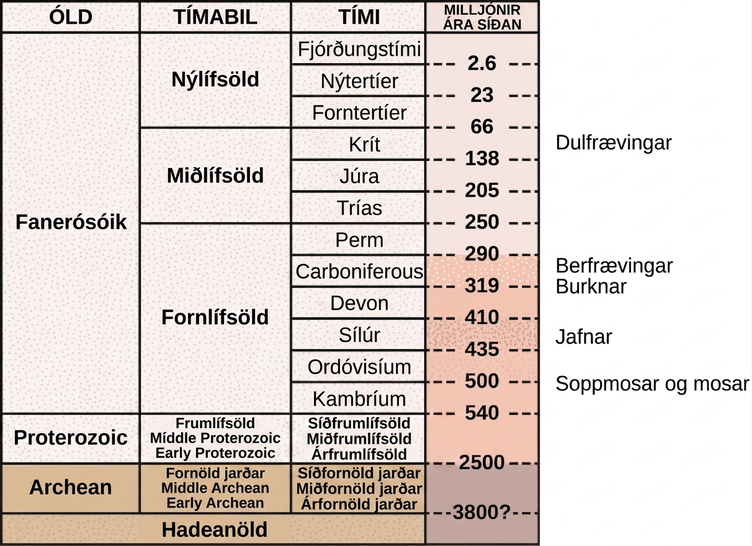
Þróun berfrævinga
Steingervingaplantan Elkinsia polymorpha, fræburkni frá devontímabilinu fyrir um 400 milljónum ára, er talin elsta fræplanta sem þekkt er til þessa. Fræburknar (Mynd 26.3) mynduðu fræ sín meðfram greinum sínum, í byggingum sem kallast bikarar og umluktu og vernduðu frævísinn, það er kvenkynliðinn og tengda vefi, sem þroskast í fræ eftir frjóvgun. Fræplöntur sem líktust nútímatrjáburknum urðu fjölmennari og fjölbreyttari í kolamýrum kolatímabilsins.

Steingervingaskráin bendir til þess að fyrstu berfrævingarnir, frumberfrævingar, hafi líklega orðið til á fornlífsöld, um miðbik devontímabilsins fyrir um 390 milljónum ára. Kolatímabilið sem fylgdi var rakt og einkenndist af risavöxnum trjáburknum. Permtímabilið þar á eftir var aftur á móti þurrt, sem gaf fræplöntum æxlunarlegt forskot vegna þess að þær voru betur lagaðar að þurrkatímum. Ginkgoales, hópur berfrævinga með aðeins eina núlifandi tegund, Ginkgo biloba, voru fyrstu berfrævingarnir sem komu fram á fyrri hluta permtímabilsins. Berfrævingar breiddust út á miðlífsöld fyrir um 240 milljónum ára, leystu burkna af hólmi í landslaginu og náðu mestri fjölbreytni sinni á þeim tíma. Júratímabilið var jafnmikið öld köngulpálma, pálmalíkra berfrævinga, og öld risaeðla. Ginkgoales og kunnuglegri barrtré settu einnig svip sinn á landslagið. Þótt dulfrævingar, eða blómplöntur, séu nú meginform plöntulífs í flestum lífbeltum ríkja berfrævingar enn í sumum vistkerfum, svo sem taigu og fjallaskógum í meiri hæð (Mynd 26.4), vegna aðlögunar sinnar að köldum og þurrum vaxtarskilyrðum.

Fræ og frjókorn sem þróunaraðlögun að þurrlendi
Gró mosa og burkna eru einlitna frumur sem eru háðar raka til að þroskast hratt í fjölfruma kynliði. Hjá fræplöntum samanstendur kvenkynliðurinn aðeins af fáeinum frumum: egginu og nokkrum stoðfrumum, þar á meðal frumu sem myndar fræhvítu og styður vöxt fósturvísisins. Eftir frjóvgun eggsins myndar tvílitna okfruman fósturvísi sem vex í grólið þegar fræið spírar. Forðavefur sem viðheldur vexti fósturvísisins og verndandi fræhjúpur veita fræjum yfirburði í þróunarlegu tilliti. Nokkur lög af hertum vef koma í veg fyrir þornun og leysa fósturvísinn undan þörf fyrir stöðugt vatnsframboð. Auk þess haldast fræ í dvala, sem þornun og hormónið abscisinsýra kalla fram, þar til vaxtarskilyrði verða hagstæð. Hvort sem fræ berast með vindi, fljóta á vatni eða eru flutt af dýrum dreifast þau yfir sífellt stærra landsvæði og forðast þannig samkeppni við móðurplöntuna.
Frjókorn (Mynd 26.5) eru karlkynliðir sem innihalda aðeins fáeinar frumur og dreifast með vindi, vatni eða frjóbera úr dýraríkinu. Öll byggingin er varin gegn þornun og getur náð til kvenlíffæranna án þess að reiða sig á vatn. Þegar frjókornið nær kvenkynliðnum myndar það frjópípu sem flytur karlkjarna að eggfrumunni. Sáðfrumur nútíma berfrævinga og allra dulfrævinga hafa ekki svipur, en hjá köngulpálmum, Ginkgo og öðrum frumstæðum berfrævingum eru sáðfrumurnar enn hreyfanlegar og nota svipur til að synda að kvenkynfrumunni. Þær berast þó til kvenkynliðsins inni í frjókorni. Frjókornið vex inn í, eða er tekið inn í, frjóvgunarhólf þar sem hreyfanlegu sáðfrumurnar losna og synda stutta vegalengd að eggi.
Þróun dulfrævinga
Þær um 200 milljónir ára sem liðu milli tilkomu berfrævinga og blómplantna gefa nokkra mynd af þeirri þróunarlegu tilraunastarfsemi sem að lokum leiddi til blóma og aldina. Dulfrævingar mynda blóm sem innihalda karllegar og/eða kvenlegar æxlunarbyggingar. Steingervingagögn (Mynd 26.6) benda til þess að blómplöntur hafi fyrst komið fram fyrir um 125 milljónum ára á fyrri hluta krítartímabilsins, seint á miðlífsöld, og hafi verið farnar að fjölbreytast hratt fyrir um 100 milljónum ára um miðbik krítartímabilsins. Eldri ummerki dulfrævinga eru fágæt. Steingerð frjókorn úr jarðlögum frá júratímabilinu hafa verið eignuð dulfrævingum. Nokkur berglög frá fyrri hluta krítartímabilsins sýna skýr för eftir laufblöð sem líkjast laufblöðum dulfrævinga. Um miðbik krítartímabilsins fyllist steingervingaskráin af gífurlegum fjölda fjölbreyttra blómplantna. Sama jarðsögutímabil markast einnig af tilkomu margra nútímahópa skordýra, sem bendir til þess að frævandi skordýr hafi gegnt lykilhlutverki í þróun blómplantna.
Ný gögn úr samanburðarlífgenafræði og forngrasafræði hafa varpað nokkru ljósi á þróun dulfrævinga. Þótt dulfrævingar hafi komið fram á eftir berfrævingum eru þeir líklega ekki komnir af forfeðrum berfrævinga. Í staðinn mynda dulfrævingar systurklæði sem þróaðist samhliða berfrævingum. Nýjungarnar tvær, blóm og aldin, sýna bætta æxlunarstefnu sem verndaði fósturvísinn og jók um leið erfðabreytileika og útbreiðslu. Engin full samstaða er nú um uppruna dulfrævinga. Forngrasafræðingar deila um hvort dulfrævingar hafi þróast úr litlum viðarkenndum runnum eða verið skyldir forfeðrum hitabeltisgrasa. Báðar skoðanir njóta stuðnings í klæðagreiningu, og svokölluð viðarkennd magnólíutilgátan, sem leggur til að snemmbúnir forfeður dulfrævinga hafi verið runnar líkir nútímamagnólíum, hefur einnig sameindalíffræðileg gögn sér til stuðnings.
Talið er að frumstæðasti núlifandi dulfrævingurinn sé Amborella trichopoda, lítil planta sem á uppruna sinn í regnskógum Nýju-Kaledóníu, eyju í Suður-Kyrrahafi. Greining á erfðamengi A. trichopoda hefur sýnt að hún er skyld öllum núlifandi blómplöntum og tilheyrir elstu staðfestu grein ættartrés dulfrævinga. Kjarnaerfðamengið sýnir merki um forna tvöföldun alls erfðamengisins. Hvatberaerfðamengið er stórt og marglitninga og inniheldur þætti úr hvatberaerfðamengjum nokkurra annarra tegunda, þar á meðal þörunga og mosa. Nokkrir aðrir hópar dulfrævinga, sem kallast grunndulfrævingar, eru taldir hafa forfeðraeinkenni vegna þess að þeir greindust snemma frá þróunartrénu. Flestum nútíma dulfrævingum er skipt í annaðhvort einkímblöðunga eða raun-tvíkímblöðunga eftir byggingu laufblaða og fósturvísa. Grunndulfrævingar, svo sem vatnaliljur, eru taldir frumstæðari vegna þess að þeir deila formfræðilegum einkennum með bæði einkímblöðungum og raun-tvíkímblöðungum.

Blóm og aldin sem þróunaraðlögun
Dulfrævingar mynda kynfrumur sínar í aðskildum líffærum sem eru venjulega í blómi. Bæði frjóvgun og þroskun fósturvísis eiga sér stað inni í líffærafræðilegri byggingu sem veitir stöðugt kerfi kynæxlunar og skýlir því að mestu fyrir sveiflum í umhverfinu. Með um 300.000 tegundir eru blómplöntur fjölbreyttasta fylking jarðar á eftir skordýrum, sem telja um 1.200.000 tegundir. Blóm eru afar fjölbreytt að stærð, lögun, lit, lykt og niðurröðun. Flest blóm hafa frjóbera sem eru í gagnkvæmum tengslum við plöntuna, og sérkenni blómanna endurspegla eðli frjóberans. Samband frjóbera og blómeinkenna er eitt skýrasta dæmið um samþróun.
Eftir frjóvgun eggsins vex frævísirinn og verður að fræi. Umlykjandi vefir egglegsins þykkna og þroskast í aldin sem verndar fræið og tryggir oft dreifingu þess yfir stórt landsvæði. Ekki þroskast öll aldin eingöngu úr egglegi; slík gervialdin þroskast úr vefjum sem liggja að eggleginu. Líkt og blóm geta aldin verið gríðarlega breytileg í útliti, stærð, lykt og bragði. Tómatar, græn paprika, maís og lárperur eru allt dæmi um aldin. Ásamt frjókornum og fræjum virka aldin einnig sem dreifingareiningar. Sum geta borist með vindi. Mörg laða að dýr sem éta aldinið og láta fræin fara í gegnum meltingarveg sinn áður en þau skila fræjunum á öðrum stað. Krókaldin eru þakin stífum, krókóttum göddum sem geta fest sig í feldi eða fatnaði og fengið far með dýri um langan veg. Krókaldinin sem festust við flauelsbuxur svissneska göngumannsins George de Mestral veittu honum innblástur að uppfinningu króka- og lykkjufestingarinnar sem hann nefndi Velcro.
Þróunartengsl
Gerð þróunartrjáa með greiningu DNA-raðsamstillinga
Allar lífverur sýna skyldleikamynstur sem má rekja til þróunarsögu þeirra. Þróunarsaga er sú fræðilega lýsing á tengslum lífvera með tilliti til forfeðra og afkomenda. Þróunartré, eins og þróunarsaga plantna sem sýnd er á Mynd 26.7, eru trjákenndar kvíslunarmyndir sem sýna þessi tengsl. Tegundir eru á endum greinanna. Hver greiningarpunktur, eða hnútur, er sá punktur þar sem ein flokkunareining, til dæmis tegund, greinist í tvær eða fleiri tegundir.
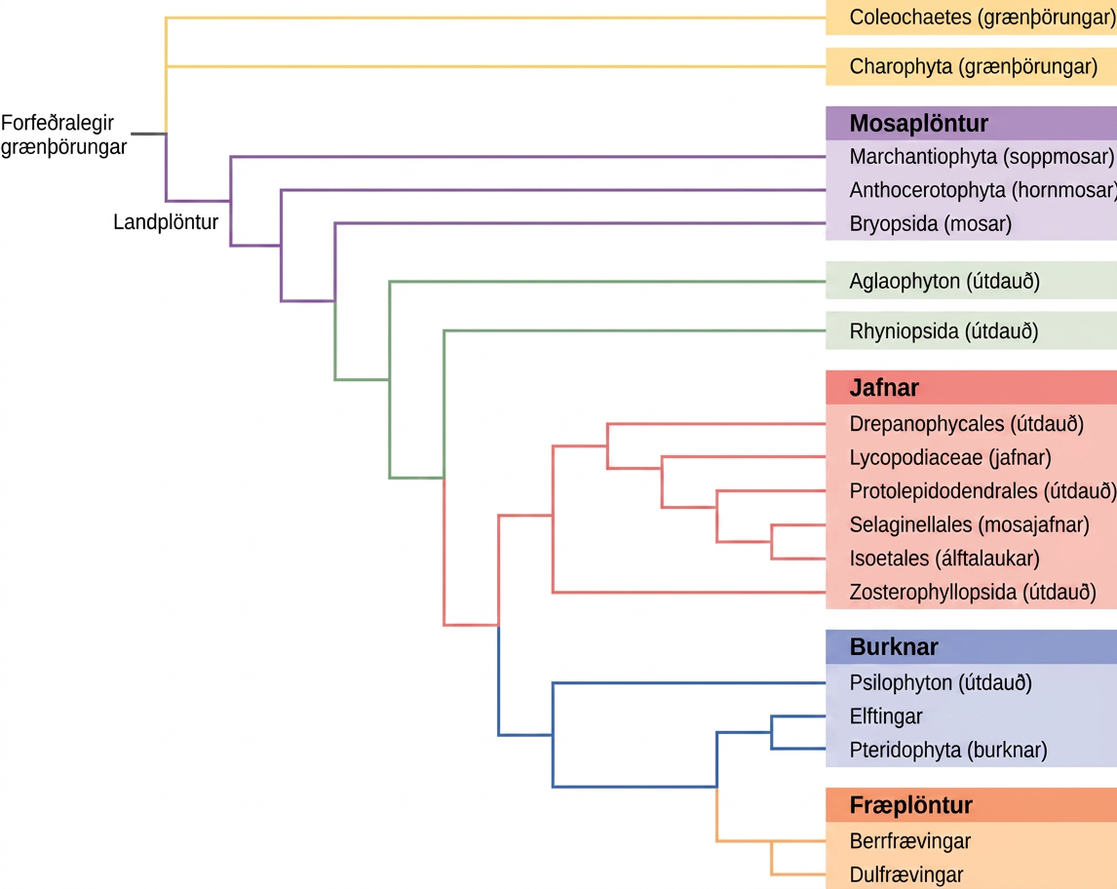
Þróunartré hafa verið byggð til að lýsa tengslum milli tegunda allt frá fyrstu skissu af tré sem birtist í verki Darwins Origin of Species. Hefðbundnar aðferðir fela í sér samanburð á samstæðum líffærafræðilegum byggingum og fósturþroska, út frá þeirri forsendu að náskyldar lífverur deili líffærafræðilegum einkennum sem koma fram við þroskun fósturvísis. Sum einkenni sem hverfa hjá fullorðnu dýri eru til staðar í fósturvísinum; til dæmis hefur snemmbúinn fósturvísir manns hala aftan við endaþarmsop, eins og allir meðlimir fylkingarinnar Chordata. Rannsóknir á steingervingum sýna millistig sem tengja forfeðraform við afkomendur þess. Margar nálganir við flokkun sem byggja eingöngu á steingervingaskránni eru þó ónákvæmar og bjóða upp á margar túlkanir. Eftir því sem verkfæri sameindalíffræði og tölvugreiningar hafa þróast á undanförnum árum hefur ný kynslóð aðferða til trjágerðar tekið á sig mynd. Lykilforsendan er sú að gen fyrir lífsnauðsynleg prótein eða RNA-byggingar, svo sem ríbósómal RNA, séu varðveitt í eðli sínu vegna þess að stökkbreytingar, breytingar á DNA-röðinni, gætu ógnað lifun lífverunnar. DNA úr örsmáum sýnum af lifandi lífverum eða steingervingum má magna með fjölliðukeðjuhvarfi (PCR) og raðgreina með því að miða á þau svæði erfðamengisins sem líklegust eru til að vera varðveitt milli tegunda. Genin sem kóða fyrir 18S rRNA litlu undireiningarinnar og plastíðgen eru oft valin til greiningar með DNA-raðsamstillingu.
Þegar raðirnar sem áhugi er á hafa fengist eru þær bornar saman við fyrirliggjandi raðir í gagnagrunnum eins og GenBank, sem National Center for Biotechnology Information heldur úti. Fjöldi tölvuverkfæra er tiltækur til að samstilla og greina raðir. Þróuð tölvugreiningarforrit ákvarða hlutfall raðasamsvörunar eða raðsamstæðu. Raðsamstæða má nota til að meta þróunarlega fjarlægð milli tveggja DNA-raða og endurspegla þann tíma sem liðinn er síðan genin aðskildust frá sameiginlegum forföður. Sameindagreining hefur gjörbreytt gerð þróunartrjáa. Í sumum tilvikum hafa fyrri niðurstöður úr formfræðilegum rannsóknum verið staðfestar, til dæmis að Amborella trichopoda sé frumstæðasti dulfrævingur sem þekktur er. Sumum hópum og tengslum hefur hins vegar verið endurraðað vegna DNA-greiningar.