35.2 Hvernig taugafrumur eiga samskipti
35.2 Hvernig taugafrumur eiga samskipti
Markmið náms
Í lok þessa hluta muntu geta gert eftirfarandi:
Öll starfsemi taugakerfisins – allt frá einföldu hreyfiviðbragði til flóknari aðgerða eins og að mynda minningu eða taka ákvörðun – krefst þess að taugafrumur hafi samskipti sín á milli. Þó að menn noti orð og líkamstjáningu til samskipta, nota taugafrumur raf- og efnaboð. Líkt og einstaklingur í nefnd, tekur ein taugafruma venjulega við og vinnur úr skilaboðum frá mörgum öðrum taugafrumum áður en hún „tekur ákvörðun“ um að senda skilaboðin áfram til annarra taugafrumna.
Flutningur taugaboðs innan taugafrumu
Til þess að taugakerfið geti starfað verða taugafrumur að geta sent og tekið á móti boðum. Þessi boð eru möguleg vegna þess að hver taugafruma hefur hlaðna frumuhimnu (spennumunur á milli innra og utanverða hliðs), og hleðsla þessarar himnu getur breyst sem svar við taugaboðefnum sem losuð eru frá öðrum taugafrumum og umhverfisáreiti. Til að skilja hvernig taugafrumur eiga samskipti verður maður fyrst að skilja grundvöll grunnlínunnar eða „hvíldarhleðslu“ himnunnar.
Hlaðnar frumuhimnur taugafrumna
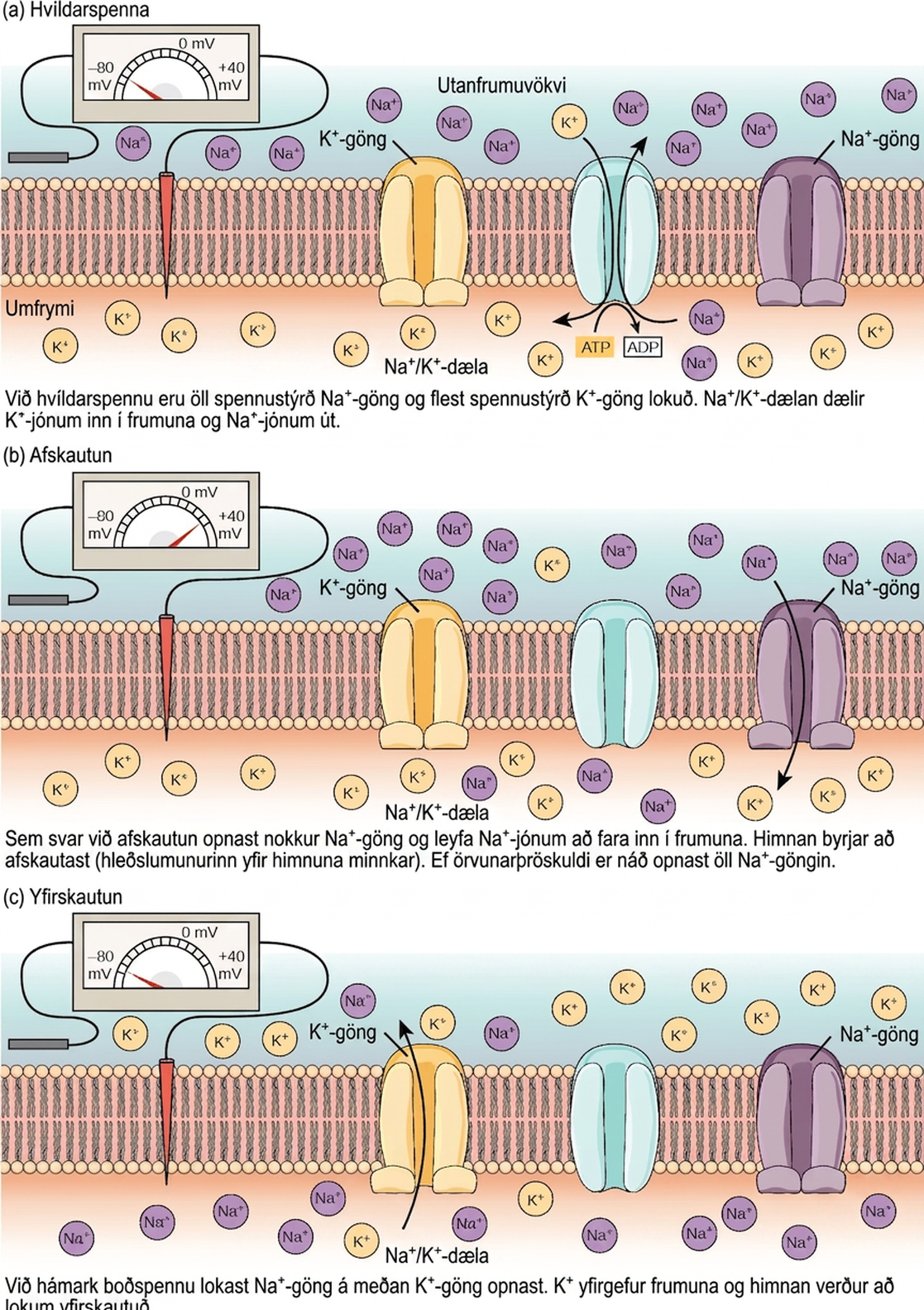
Tvöfalda lípíðlagið sem umlykur taugafrumu er ógegndræpt fyrir hlöðnum sameindum eða jónum. Til að komast inn eða út úr taugafrumunni verða jónir að fara í gegnum sérstök prótein sem kallast jónagöng og ná í gegnum himnuna. Jónagöng hafa mismunandi stillingar: opin, lokuð og óvirk, eins og sýnt er á mynd 35.9. Sum jónagöng þarf að virkja til að þau opnist og hleypi jónum inn eða út úr frumunni. Þessi jónagöng eru næm fyrir umhverfinu og geta breytt lögun sinni í samræmi við það. Jónagöng sem breyta byggingu sinni sem svar við spennubreytingum kallast spennustýrð jónagöng. Spennustýrð jónagöng stjórna hlutfallslegum styrk mismunandi jóna innan og utan frumunnar. Munurinn á heildarhleðslu milli innra og utanverða hliðs frumunnar kallast himnuspenna.

Tengill í námsefni
Þetta myndband fjallar um grundvöll hvíldarhimnuspennu.
Hvíldarhimnuspenna
Taugafruma í hvíld er neikvætt hlaðin: innanverða hlið frumu er um það bil 70 millivoltum neikvæðara en utanverða hliðð (−70 mV, athugið að þessi tala er breytileg eftir tegund taugafrumu og dýrategund). Þessi spenna kallast hvíldarhimnuspenna; hún orsakast af mismun á styrk jóna innan og utan frumunnar. Ef himnan væri jafn gegndræp fyrir öllum jónum myndi hver tegund jóna flæða yfir himnuna og kerfið næði jafnvægi. Þar sem jónir geta ekki einfaldlega farið yfir himnuna að vild, er mismunandi styrkur nokkurra jóna innan og utan frumunnar, eins og sýnt er í töflu 35.1. Mismunurinn á fjölda jákvætt hlaðinna kalíumjóna (K+) innan og utan frumunnar ræður mestu um hvíldarhimnuspennunnar (mynd 35.10). Þegar himnan er í hvíld safnast K+-jónir upp inni í frumunni vegna virkni Na+/K+-dælunnar, sem knýr báðar jónir gegn styrkfallanda sínum. Neikvæða hvíldarspennan myndast og viðhelst með því að auka styrk katjóna utan frumunnar (í utanfrumuvökvanum) miðað við innan frumunnar (í umfryminu). Neikvæða hleðslan inni í frumunni myndast vegna þess að frumuhimnan er gegndræpari fyrir flæði kalíumjóna en natríumjóna. Í taugafrumum er kalíumjónum haldið í háum styrk inni í frumunni á meðan natríumjónum er haldið í háum styrk utan frumunnar. Fruman býr yfir kalíum- og natríumlekagöngum sem leyfa þessum tveimur katjónum að flæða undan styrkfallanda sínum. Hins vegar hafa taugafrumur mun fleiri kalíumlekagöng en natríumlekagöng. Þess vegna flæðir kalíum út úr frumunni á mun meiri hraða en natríum lekur inn. Þar sem fleiri katjónir yfirgefa frumuna en koma inn, veldur þetta því að innanverða hlið frumunnar verður neikvætt hlaðið miðað við utanverða hliðð. Virkni natríum-kalíumdælunnar hjálpar til við að viðhalda hvíldarhimnuspennunni þegar hún hefur myndast. Munið að natríum-kalíumdælur flytja tvær K+-jónir inn í frumuna en fjarlægja þrjár Na+-jónir fyrir hvert ATP sem er notað. Þar sem fleiri katjónir eru fluttar út úr frumunni en teknar inn, helst innanverða hlið frumunnar neikvætt hlaðið miðað við utanfrumuvökvann. Þess má geta að klóríðjónir (Cl–) hafa tilhneigingu til að safnast upp utan frumunnar vegna þess að neikvætt hlaðin prótein í umfryminu hrinda þeim frá sér.
| Jón | Styrkur utan frumu (mM) | Styrkur innan frumu (mM) | Hlutfall utan/innan |
|---|---|---|---|
| 145 | 12 | 12 | |
| 4 | 155 | 0.026 | |
| 120 | 4 | 30 | |
| Lífrænar anjónir (A−) | — | 100 |
Boðspenna
Taugafruma getur tekið við boðum frá öðrum taugafrumum og, ef þessi boð eru nógu sterk, sent merkið til næstu taugafrumna. Flutningur boða á milli taugafrumna fer almennt fram með efni sem kallast taugaboðefni. Flutningur boða innan taugafrumu (frá griplu til símaenda) fer fram með stuttri umsnúningu á hvíldarhimnuspennunnar sem kallast boðspenna. Þegar taugaboðefnissameindir bindast viðtökum sem staðsettir eru á griplum taugafrumu opnast jónagöng. Á örvandi taugamótum leyfir þessi opnun jákvæðum jónum að fara inn í taugafrumuna og leiðir til afskautunar himnunnar – minnkunar á spennumun milli innra og utanverða hliðs taugafrumunnar. Áreiti frá skynfrumu eða annarri taugafrumu afskautar marktaugafrumuna að þröskuldsspennu hennar (-55 mV). Na+-göng í símahól opnast og leyfa jákvæðum jónum að fara inn í frumuna (mynd 35.10 og mynd 35.11). Þegar natríumgöngin opnast afskautast taugafruman alveg í himnuspennu upp á um +40 mV. Boðspennur eru taldar „allt-eða-ekkert“ atburður, þar sem taugafruman afskautast alltaf að fullu þegar þröskuldsspennu er náð. Þegar afskautun er lokið verður fruman nú að „endurstilla“ himnuspennu sína aftur í hvíldarspennu. Til að ná þessu fram lokast Na+-göngin og geta ekki opnast aftur strax. Þetta markar upphaf ónæmistíma taugafrumunnar, þar sem hún getur ekki myndað aðra boðspennu vegna þess að natríumgöng hennar opnast ekki. Á sama tíma opnast spennustýrð K+-göng sem leyfa K+ að yfirgefa frumuna. Þegar K+-jónir yfirgefa frumuna verður himnuspennan aftur neikvæð og endurskautast. Flæði K+ út úr frumunni heldur í raun áfram í stuttan tíma eftir að hvíldarspennu er náð, og himnan yfirskautast, þannig að himnuspennan verður neikvæðari en venjuleg hvíldarspenna frumunnar. Þetta er afleiðing hægrar lokunar K+-ganganna. Á þessum tímapunkti snúa natríumgöngin aftur í hvíldarástand sitt, sem þýðir að þau eru tilbúin að opnast aftur ef himnuspennan fer aftur yfir þröskuldsspennuna. Að lokum lokast öll K+-göng og fruman snýr aftur í hvíldarspennu sína.
Myndræn tengsl
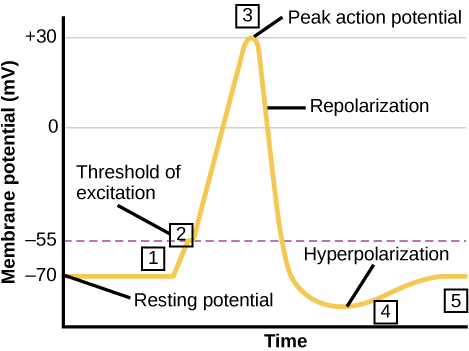
Kalíumgangahindrar, eins og amíódarón og prókaínamíð, sem eru notaðir til að meðhöndla óeðlilega rafvirkni í hjarta, kallað hjartsláttartruflanir, hindra flæði K+ í gegnum spennustýrð K+-göng. Hvaða hluta boðspennunnar myndir þú búast við að kalíumgöng hafi áhrif á?
Kalíumgangahindrar hægja á endurskautunarfasanum en hafa engin áhrif á afskautun.

Tengill í námsefni
Þetta myndband sýnir yfirlit yfir boðspennu.
Mýelín og útbreiðsla boðspennu
Til þess að boðspenna geti miðlað upplýsingum til annarrar taugafrumu verður hún að ferðast eftir símanum og ná til símaendanna þar sem hún getur komið af stað losun taugaboðefna. Hraði leiðni boðspennu eftir síma verður fyrir áhrifum af bæði þvermáli símans og viðnámi símans gegn straumleka. Mýelín virkar sem einangrari sem kemur í veg fyrir að straumur fari út úr símanum; þetta eykur hraða boðspennuleiðni. Í afmýelínunarsjúkdómum eins og heila- og mænusiggi (MS) hægist á leiðni boðspennu vegna þess að straumur lekur frá svæðum á símanum sem áður voru einangruð. Ranvierskorarnar, sem sýnd eru á mynd 35.13, eru rof í mýelínslíðrinu eftir endilöngum símanum. Þessi ómýelíniseruðu svæði eru um einn míkrómetri að lengd og innihalda spennustýrð Na+ og K+-göng. Flæði jóna í gegnum þessi göng, sérstaklega Na+-göngin, endurmyndar boðspennuna aftur og aftur eftir símanum. Þetta „stökk“ boðspennunnar frá einni Ranvierskor til þeirrar næstu kallast stökkleiðni. Ef Ranvierskor væru ekki til staðar eftir símanum myndi boðspennan breiðast mjög hægt út þar sem Na+ og K+-göng þyrftu stöðugt að endurmynda boðspennur á hverjum punkti eftir símanum í stað þess að gera það á tilteknum punktum. Ranvierskor spara einnig orku fyrir taugafrumuna þar sem göngin þurfa aðeins að vera til staðar við Ranvierskorarnar en ekki eftir öllum símanum.

Boðflutningur um taugamót
Taugamótin eða „bilið“ er staðurinn þar sem upplýsingar eru fluttar frá einni taugafrumu til annarrar. Taugamót myndast venjulega á milli símaenda og griplutinda, en það er ekki algilt. Einnig eru til taugamót milli síma og síma, griplu og griplu, og síma og frumubols. Taugafruman sem sendir boðið kallast fyrirmótafruma og taugafruman sem tekur við boðinu kallast eftirmótafruma. Athugið að þessi heiti eru afstæð miðað við tiltekin taugamót – flestar taugafrumur eru bæði fyrirmóta- og eftirmótafrumur. Það eru tvær gerðir taugamóta: efnataugamót og raftaugamót.
Efnataugamót
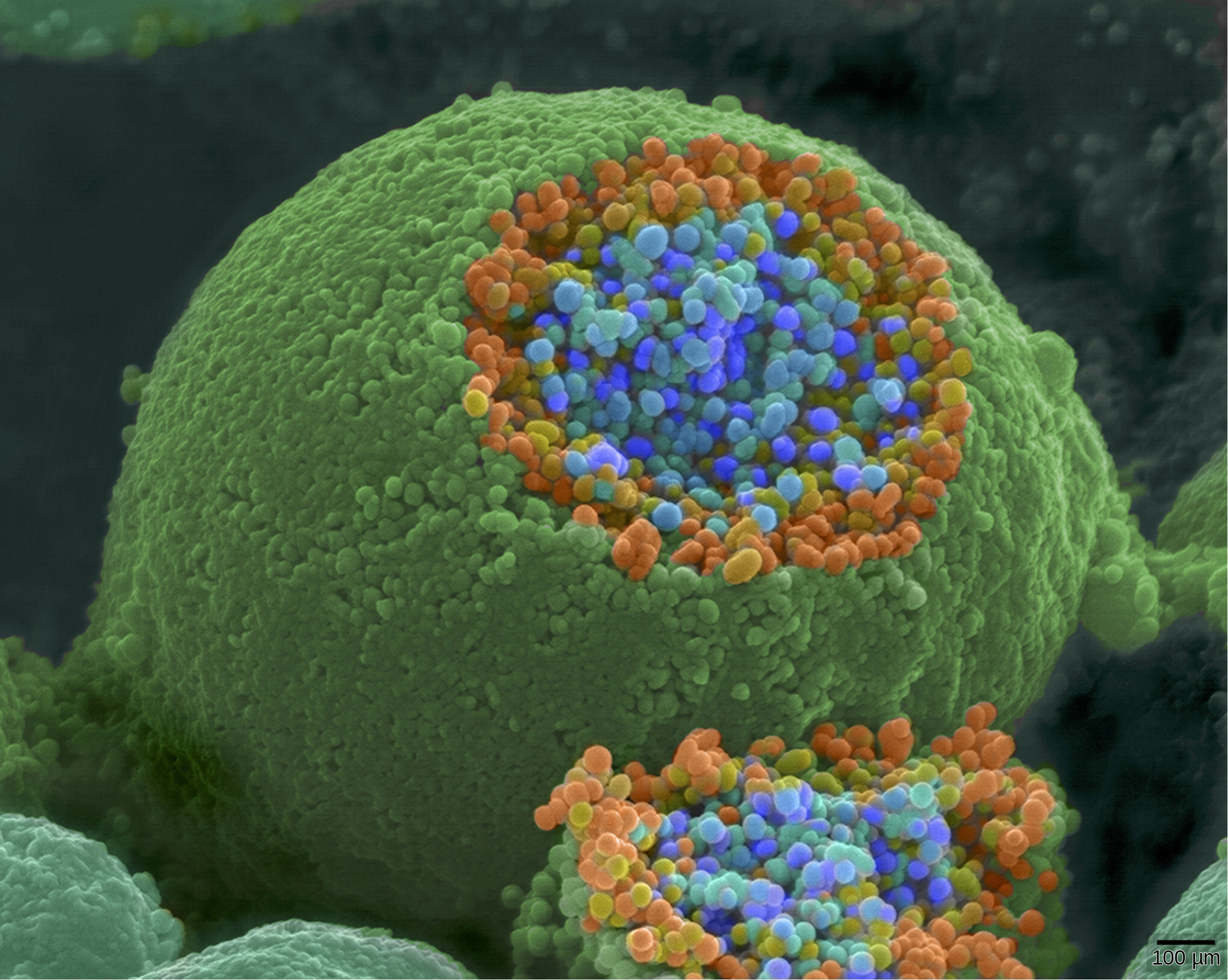
Þegar boðspenna nær til símaenda afskautar hún himnuna og opnar spennustýrð Na+-göng. Na+-jónir streyma inn í frumuna og afskauta fyrirmótahimnuna enn frekar. Þessi afskautun veldur því að spennustýrð Ca2+-göng opnast. Kalsíumjónir sem fara inn í frumuna koma af stað boðferli sem veldur því að litlar himnubundnar bólur, kallaðar taugamótabólur, sem innihalda taugaboðefnissameindir, renna saman við fyrirmótahimnuna. Taugabólur eru sýndar á mynd 35.14, sem er mynd úr skönnunarrafeindasmásjá.
Samruni bólu við fyrirmótahimnuna veldur því að taugaboðefni losnar út í taugamótaglufuna, utanfrumurýmið milli fyrirmóta- og eftirmótahimnanna, eins og sýnt er á mynd 35.15. Taugaboðefnið flæðir yfir taugamótaglufuna og binst viðtakapróteinum á eftirmótahimnunni.

Binding tiltekins taugaboðefnis veldur því að ákveðin jónagöng, í þessu tilviki bindilstýrð göng, á eftirmótahimnunni opnast. Taugaboðefni geta annaðhvort haft örvandi eða hamlandi áhrif á eftirmótahimnuna. Til dæmis, þegar asetýlkólín er losað á taugamótum milli taugar og vöðva (kallað taugavöðvamót) af fyrirmótafrumu, veldur það því að Na+-göng eftirmótafrumu opnast. Na+ fer inn í eftirmótafrumuna og veldur því að eftirmótahimnan afskautast. Þessi afskautun kallast örvandi eftirmótaspenna (EPSP) og gerir það líklegra að eftirmótafruman sendi boðspennu. Losun taugaboðefnis á hamlandi taugamótum veldur hamlandi eftirmótaspennu (IPSP-boð), sem er yfirskautun á eftirmótahimnunni. Til dæmis, þegar taugaboðefnið GABA (gamma-amínósmjörsýra) er losað frá fyrirmótafrumu, binst það og opnar Cl−-göng. Cl−-jónir fara inn í frumuna og yfirskauta himnuna, sem gerir það ólíklegra að taugafruman sendi boðspennu.
Þegar taugaboðflutningur hefur átt sér stað verður að fjarlægja taugaboðefnið úr taugamótaglufunni svo eftirmótahimnan geti „endurstillt sig“ og verið tilbúin að taka við öðru boði. Þetta er hægt að gera á þrjá vegu: taugaboðefnið getur flætt burt frá taugamótaglufunni, það getur verið brotið niður af ensímum í taugamótaglufunni, eða það getur verið endurunnið (stundum kallað endurupptaka) af fyrirmótafrumunni. Nokkur lyf virka á þetta skref taugaboðflutnings. Til dæmis virka sum lyf sem gefin eru Alzheimer-sjúklingum með því að hamla asetýlkólínesterasa, ensímið sem brýtur niður asetýlkólín. Þessi hömlun á ensíminu eykur í raun taugaboðflutning á taugamótum sem losa asetýlkólín. Þegar asetýlkólín hefur verið losað helst það í glufunni og getur stöðugt bundist og losnað frá viðtökum á eftirmótafrumu.
| Taugaboðefni | Dæmi | Staðsetning |
|---|---|---|
| Asetýlkólín | — | CNS og/eða PNS |
| Lífmyndað amín | Dópamín, serótónín, noradrenalín | CNS og/eða PNS |
| Amínósýra | Glýsín, glútamat, aspartat, gamma-amínósmjörsýra | CNS |
| Taugapeptíð | P-efni, endorfín | CNS og/eða PNS |
Raftaugamót
Þótt raftaugamót séu færri en efnataugamót finnast þau í öllum taugakerfum og gegna mikilvægum og einstökum hlutverkum. Máti taugaboðflutnings í raftaugamótum er nokkuð frábrugðinn því sem gerist í efnataugamótum. Í raftaugamótum eru fyrirmóta- og eftirmótahimnurnar mjög þétt saman og eru í raun tengdar efnislega með gangapróteinum sem mynda gatatengi. Gatatengi leyfa straumi að fara beint frá einni frumu til þeirrar næstu. Auk jónanna sem bera þennan straum geta aðrar sameindir, eins og ATP, flætt í gegnum stór op gatatengjanna.
Það er lykilmunur á efnataugamótum og raftaugamótum. Vegna þess að efnataugamót treysta á losun taugaboðefnissameinda úr taugamótabólum til að koma boðum sínum áfram, er um það bil einnar millisekúndu töf frá því að boðspenna nær til fyrirmótaenda og þar til taugaboðefnið leiðir til opnunar jónaganga í eftirmótafrumu. Að auki eru þessi boðskipti einátta. Boðskipti í raftaugamótum eru hins vegar nánast samstundis (sem er mikilvægt fyrir taugamót sem koma að lykilviðbrögðum) og sum raftaugamót eru tvíátta. Raftaugamót eru einnig áreiðanlegri þar sem ólíklegra er að þau stíflist, og þau eru mikilvæg fyrir samstillingu rafvirkni í hópi taugafrumna. Til dæmis er talið að raftaugamót í stúku (thalamus) stjórni hægbylgjusvefni og truflun á þessum taugamótum getur valdið krampaflogum.
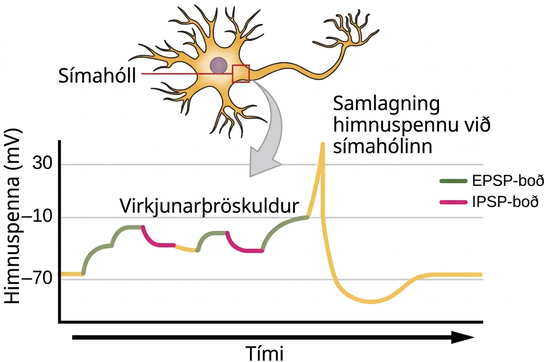
Samlagning boða
Stundum er eitt EPSP nógu sterkt til að framkalla boðspennu í eftirmótafrumu, en oft þurfa mörg fyrirmótainntök að skapa EPSP-boð um svipað leyti til að eftirmótafruman afskautist nægilega til að senda boðspennu. Þetta ferli kallast samlagning og á sér stað við símahólinn, eins og sýnt er á mynd 35.16. Að auki hefur ein taugafruma oft inntök frá mörgum fyrirmótafrumum – sumum örvandi og sumum hamlandi – svo IPSP-boð geta eytt út EPSP-boð og öfugt. Það er nettóbreytingin á spennu eftirmótahimnunnar sem ákvarðar hvort eftirmótafruman hafi náð þröskuldi örvunar sem þarf til að senda boðspennu. Saman virka samlagning taugamóta og þröskuldur örvunar sem sía þannig að handahófskenndur „suð“ í kerfinu berist ekki áfram sem mikilvægar upplýsingar.
Tenging við daglegt líf
Viðmót heila og tölvu
Hreyfitaugungahrörnun (ALS, einnig kallað Lou Gehrig-sjúkdómurinn) er taugasjúkdómur sem einkennist af hrörnun hreyfitaugunga sem stjórna viljastýrðum hreyfingum. Sjúkdómurinn byrjar með vöðvaslappleika og skorti á samhæfingu og eyðileggur að lokum taugafrumurnar sem stjórna tali, öndun og kyngingu; á endanum getur sjúkdómurinn leitt til lömunar. Á þeim tímapunkti þurfa sjúklingar aðstoð véla til að geta andað og tjáð sig. Nokkrar sérstakar tæknilausnir hafa verið þróaðar til að gera „innilokuðum“ (e. locked-in) sjúklingum kleift að eiga samskipti við umheiminn. Ein tækni gerir sjúklingum til dæmis kleift að skrifa setningar með því að hreyfa kinnina. Þessar setningar geta síðan verið lesnar upphátt af tölvu.
Tiltölulega ný rannsóknarstefna til að hjálpa lömuðum sjúklingum, þar á meðal þeim með ALS, við að tjá sig og halda í nokkurt sjálfstæði kallast heila-tölvuviðmót (BCI) og er sýnd á mynd 35.17. Þessi tækni hljómar eins og eitthvað úr vísindaskáldskap: hún gerir lömuðum sjúklingum kleift að stjórna tölvu eingöngu með hugsunum sínum. Það eru til nokkrar gerðir af BCI. Sumar gerðir nota EEG upptökur frá rafskautum sem eru límd á höfuðkúpuna. Þessar upptökur innihalda upplýsingar frá stórum hópum taugafrumna sem tölva getur afkóðað. Aðrar gerðir af BCI krefjast ígræðslu á fylki rafskauta, sem er minna en frímerki, í handleggs- og handarsvæði hreyfibarkarins. Þessi tegund af BCI, þó hún sé meira inngrip, er mjög öflug þar sem hvert rafskaut getur skráð raunverulegar boðspennur frá einni eða fleiri taugafrumum. Þessi merki eru síðan send í tölvu, sem hefur verið þjálfuð til að afkóða merkið og mata það í verkfæri – eins og bendil á tölvuskjá. Þetta þýðir að sjúklingur með ALS getur notað tölvupóst, lesið á netinu og átt samskipti við aðra með því að hugsa um að hreyfa höndina eða handlegginn (jafnvel þótt lamaði sjúklingurinn geti ekki framkvæmt þá líkamlegu hreyfingu). Nýlegar framfarir hafa gert lömuðum sjúklingi í innilokunarástandi, sem fékk heilablóðfall fyrir 15 árum, kleift að stjórna vélmennaarm og jafnvel gefa sjálfri sér kaffi með því að nota BCI-tækni.
Þrátt fyrir ótrúlegar framfarir í BCI-tækni, hefur hún einnig takmarkanir. Tæknin getur krafist margra klukkustunda þjálfunar og langra tímabila af mikilli einbeitingu fyrir sjúklinginn; hún getur einnig krafist heilaskurðaðgerðar til að græða tækin í.

Tengill í námsefni
Horfðu á þetta myndband þar sem lömuð kona notar heilastýrðan vélmennaarm til að bera drykk að munni sér, ásamt öðrum myndum af heila-tölvuviðmóttækni í verki.
Mótanleiki taugamóta
Taugamót eru ekki stöðugar byggingar. Þau geta veikst eða styrkst. Þau geta rofnað og ný taugamót geta myndast. Mótanleiki taugamóta gerir þessar breytingar mögulegar, en þær eru allar nauðsynlegar fyrir virkt taugakerfi. Í raun er mótanleiki taugamóta grundvöllur náms og minnis. Tvö ferli sérstaklega, langtímaefling (LTP) og langtímabæling (LTD), eru mikilvæg form mótanleika taugamóta sem eiga sér stað í taugamótum í dreka (hippocampus), heilasvæði sem tekur þátt í geymslu minninga.
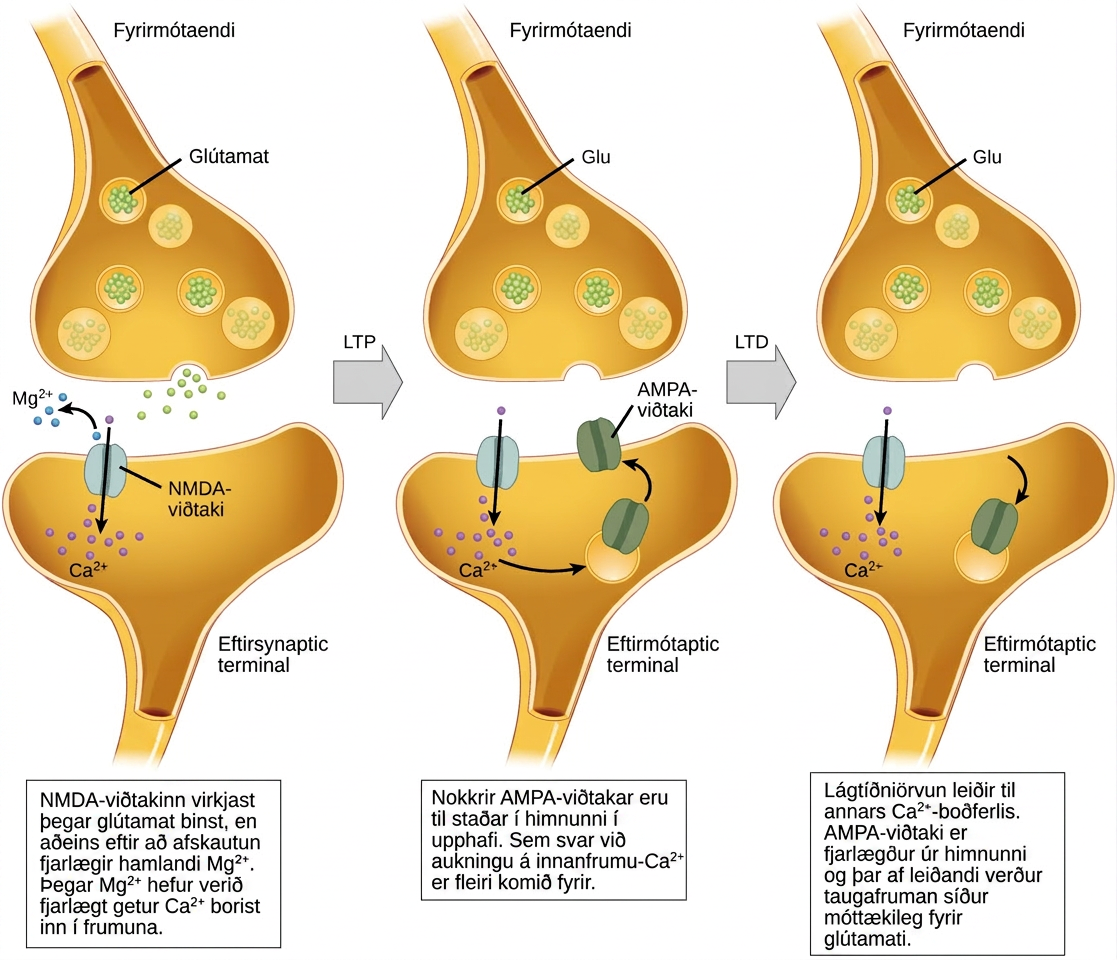
Langtímaefling (LTP)
Langtímaefling (LTP) er viðvarandi styrking á tengingu taugamóta. LTP byggir á reglu Hebbs: frumur sem senda boð saman, tengjast saman. Það eru ýmis ferli, ekkert þeirra fullskilið, á bak við styrkingu taugamóta sem sést við LTP. Eitt þekkt ferli felur í sér tegund af glútamatviðtaka á eftirmótafrumu, kallaða NMDA (N-metýl-D-aspartat) viðtaka, sem sýndir eru á mynd 35.18. Þessir viðtakar eru venjulega blokkaðir af magnesíumjónum; hins vegar, þegar eftirmótataugafruman er afskautuð af mörgum boðum frá fyrirmótafrumum í snöggri röð (annaðhvort frá einni taugafrumu eða mörgum), þvingast magnesíumjónirnar út og leyfa Ca2+-jónum að fara inn í eftirmótafrumuna. Næst koma Ca2+ jónir inn í frumuna og koma af stað boðferli sem veldur því að önnur tegund glútamatviðtaka, kölluð AMPA (α-amínó-3-hýdroxý-5-metýl-4-ísóxazólprópíónsýra) viðtakar, eru settir inn í eftirmótahimnuna, þar sem virkjaðir AMPA-viðtakar leyfa jákvæðum jónum að fara inn í frumuna. Svo, næst þegar glútamat losnar frá fyrirmótahimnunni, mun það hafa meiri örvandi áhrif (EPSP) á eftirmótafrumuna vegna þess að binding glútamats við þessa AMPA-viðtaka mun hleypa fleiri jákvæðum jónum inn í frumuna. Innsetning fleiri AMPA-viðtaka styrkir taugamótin og þýðir að líklegra er að eftirmótataugafruman sendi boð sem svar við losun taugaboðefna frá fyrirmótafrumu. Sum fíkniefni nýta sér LTP ferlið og þessi styrking taugamóta getur leitt til fíknar.
Langtímabæling (LTD)
Langtímabæling (LTD) er í raun andstæðan við LTP: hún er langtímaveiking á tengingu taugamóta. Eitt ferli sem vitað er að veldur LTD felur einnig í sér AMPA-viðtaka. Í þessu tilviki kemur kalsíum sem fer inn í gegnum NMDA-viðtaka af stað öðru boðferli, sem leiðir til þess að AMPA-viðtakar eru fjarlægðir úr eftirmótahimnunni, eins og sýnt er á mynd 35.18. Fækkun AMPA-viðtaka í himnunni gerir eftirmótataugafrumuna minna móttækilega fyrir glútamati sem losnar frá fyrirmótataugafrumunni. Þó að það kunni að virðast órökrétt, getur LTD verið jafn mikilvæg fyrir nám og minni og LTP. Veiking og grisjun ónotaðra taugamóta gerir það að verkum að ómikilvægar tengingar tapast og gerir taugamótin sem hafa gengist undir LTP þeim mun sterkari í samanburði.