14.5 Eftirmyndun DNA í heilkjörnungum
14.5 Eftirmyndun DNA í heilkjörnungum
Námsmarkmið
Í lok þessa hluta muntu geta gert eftirfarandi:
- Rætt líkindi og mun á eftirmyndun DNA í heilkjörnungum og dreifkjörnungum
- Lýst hlutverki telómerasa í eftirmyndun DNA
Erfðamengi heilkjörnunga eru mun flóknari og stærri en erfðamengi dreifkjörnunga. Heilkjörnungar hafa einnig marga ólíka línulega litninga. Erfðamengi mannsins hefur 3 milljarða basapara í hverju einlitnu litningasetti og 6 milljarðar basapara eru eftirmyndaðir í S-fasa frumuhringsins. Margir upphafsstaðir eftirmyndunar eru á hverjum litningi heilkjörnunga; menn geta haft allt að 100.000 upphafsstaði eftirmyndunar í erfðamenginu. Hraði eftirmyndunar er um það bil 100 kirni á sekúndu, mun hægari en eftirmyndun hjá dreifkjörnungum. Í gersveppum, sem eru heilkjörnungar, finnast sérstakar raðir á litningunum sem kallast sjálfstætt eftirmyndandi raðir (ARS). Þær samsvara upphafsstað eftirmyndunar í E. coli.
Fjöldi DNA-pólýmerasa í heilkjörnungum er mun meiri en í dreifkjörnungum: 14 eru þekktir og þar af eru fimm vel rannsakaðir og vitað er að þeir gegna meginhlutverki við eftirmyndun. Þeir eru þekktir sem pol α, pol β, pol γ, pol δ og pol ε.
Grundvallarskref eftirmyndunar eru þau sömu og í dreifkjörnungum. Áður en eftirmyndun getur hafist verður DNA að vera aðgengilegt sem mót. DNA heilkjörnunga er bundið basískum prótínum sem kallast histón og myndar þannig byggingar sem kallast litnisagnir. Fjarlægja þarf histón og setja þau síðan aftur á sinn stað meðan á eftirmyndun stendur, sem skýrir að hluta lægri eftirmyndunarhraða í heilkjörnungum. Litni, flóki DNA og prótína, getur tekið efnafræðilegum breytingum þannig að DNA losni frá prótínunum eða verði aðgengilegt ensímum eftirmyndunarvélar DNA. Við upphafsstað eftirmyndunar myndast foreftirmyndunarflóki ásamt öðrum upphafsprótínum. Helíkasi og önnur prótín eru síðan kölluð til til að hefja eftirmyndunarferlið (tafla 14.2).
| Eiginleiki | Dreifkjörnungar | Heilkjörnungar |
|---|---|---|
| Upphafsstaðir eftirmyndunar | Einn | Margir |
| Hraði eftirmyndunar | 1000 kirni/s | 50 til 100 kirni/s |
| Gerðir DNA-pólýmerasa | 5 | 14 |
| Telómerasi | Ekki til staðar | Til staðar |
| Fjarlæging RNA-vísa | DNA pol I | RNasi H |
| Lenging DNA-þráða | DNA pol III | pol α, pol δ, pol ε |
| Renniklemma | Renniklemma |
Helíkasi sem notar orku frá ATP-vatnsrofi opnar DNA-gorminn. Eftirmyndunarkvíslar myndast við hvern upphafsstað eftirmyndunar þegar DNA vindur ofan af sér. Opnun tvöfalda gormsins veldur yfirvindingu, eða ofurvindingu, í DNA fyrir framan eftirmyndunarkvíslina. Tópóísómerasar leysa úr þessari spennu. Vísar eru myndaðir af ensíminu prímasa og DNA pol getur hafið nýmyndun með því að nota vísinn. Þrír megin-DNA-pólýmerasar koma síðan við sögu: α, δ og ε. DNA pol α bætir stuttum DNA-bút, 20 til 30 kirna löngum, við RNA-vísinn á báðum þráðum og afhendir síðan verkið öðrum pólýmerasa. Leiðandi strengurinn er nýmyndaður samfellt af ensíminu pol ε en seinni strengurinn er nýmyndaður af pol δ. Renniklemmuprótín sem kallast PCNA (proliferating cell nuclear antigen) heldur DNA pol á sínum stað svo það renni ekki af DNA. Þegar pol δ rekst á RNA-vísinn á seinni strengnum losar það vísinn frá DNA-mótinu. Losaði RNA-vísirinn er síðan fjarlægður af RNasa H, einnig kölluðum flipaendónúkleasa, og DNA-kirni koma í staðinn. Okazaki-bútarnir á seinni strengnum eru tengdir saman eftir að RNA-vísunum hefur verið skipt út fyrir DNA. Bilin sem eftir standa eru lokuð af DNA-lígasa, sem myndar fosfódíestertengið.
Eftirmyndun telómera
Ólíkt litningum dreifkjörnunga eru litningar heilkjörnunga línulegir. Eins og þú hefur lært getur ensímið DNA pol aðeins bætt við kirnum í 5′ til 3′ stefnu. Á leiðandi strengnum heldur nýmyndun áfram þar til enda litningsins er náð. Á seinni strengnum er DNA nýmyndað í stuttum bútum sem hver um sig hefst með sérstökum vísi. Þegar eftirmyndunarkvíslin nær enda línulega litningsins er engin leið til að skipta út vísinum á 5′ enda seinni strengsins. DNA á endum litningsins verður því eftir óparað og með tímanum geta þessir endar, sem kallast telómerar, styst smám saman eftir því sem frumur halda áfram að skipta sér.
Telómerar samanstanda af endurteknum röðum sem kóða ekki fyrir neinu tilteknu geni. Í mönnum er sex basapara röðin TTAGGG endurtekin 100 til 1000 sinnum á telómerasvæðunum. Að vissu leyti vernda telómerar genin gegn því að þau glatist þegar frumur halda áfram að skipta sér. Telómerum er bætt við enda litninganna með sérstöku ensími, telómerasa (mynd 14.16), og uppgötvun þess hjálpaði til við að skýra hvernig þessum endurteknu litningaendum er viðhaldið. Telómerasaensímið inniheldur hvatahluta og innbyggt RNA-mót. Það festist við enda litningsins og DNA-kirnum sem eru mótsvarandi RNA-mótinu er bætt við 3′ enda DNA-strengsins. Þegar 3′ endi móts seinni strengsins hefur verið lengdur nægilega getur DNA-pólýmerasi bætt við kirnum sem eru mótsvarandi endum litninganna. Þannig eru endar litninganna eftirmyndaðir.
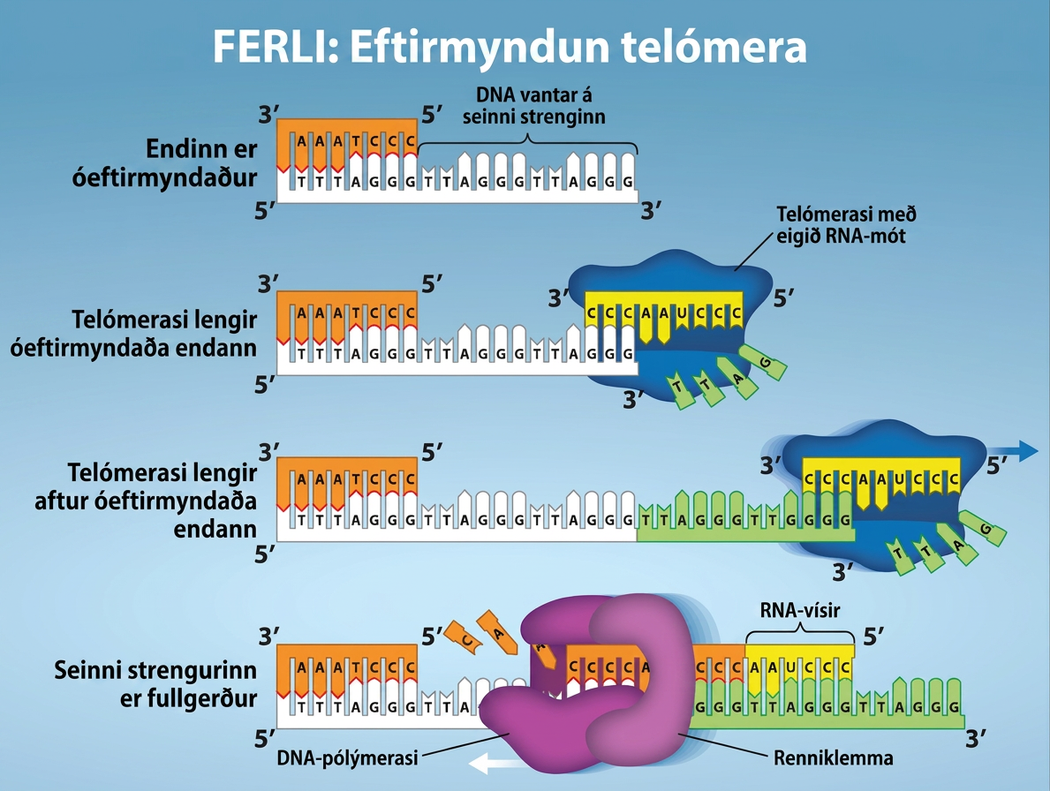
Telómerasi er yfirleitt virkur í kynstofnfrumum og fullorðinsstofnfrumum. Hann er ekki virkur í líkamsfrumum fullorðinna. Fyrir uppgötvun sína á telómerasa og virkni hans fengu Elizabeth Blackburn, Carol W. Greider og Jack W. Szostak (mynd 14.16) Nóbelsverðlaunin í lífeðlisfræði eða læknisfræði árið 2009. Síðari rannsóknir á HeLa-frumum, fengnum frá Henrietta Lacks, staðfestu að telómerasi er til staðar í frumum manna. Árið 2001 fundu vísindamenn, þar á meðal Diane L. Wright, að telómerasi er nauðsynlegur til að frumur í fósturvísum manna geti fjölgað sér hratt.

Telómerasi og öldrun
Frumur sem ganga í gegnum frumuskiptingu fá sífellt styttri telómera vegna þess að flestar líkamsfrumur framleiða ekki telómerasa. Þetta þýðir í raun að stytting telómera tengist öldrun. Með tilkomu nútímalæknisfræði, fyrirbyggjandi heilbrigðisþjónustu og heilbrigðari lífsstíls hafa lífslíkur manna aukist og sífellt meiri eftirspurn er eftir því að fólk líti yngra út og hafi betri lífsgæði þegar það eldist.
Árið 2010 komust vísindamenn að því að telómerasi getur snúið við sumum aldurstengdum kvillum í músum. Þetta gæti nýst í endurnýjunarlæknisfræði.2 Mýs sem skorti telómerasa voru notaðar í þessum rannsóknum; þær sýndu vefjarýrnun, eyðingu stofnfrumna, bilun í líffærakerfum og skerta svörun við vefjaskaða. Endurvirkjun telómerasa í þessum músum olli lengingu telómera, dró úr DNA-skemmdum, sneri við taugahrörnun og bætti virkni eistna, milta og þarma. Þannig gæti endurvirkjun telómerasa nýst við meðferð aldurstengdra sjúkdóma í mönnum.
Krabbamein einkennist af stjórnlausri frumuskiptingu óeðlilegra frumna. Frumurnar safna stökkbreytingum, fjölga sér stjórnlaust og geta flust til ólíkra hluta líkamans í ferli sem kallast meinvörp. Vísindamenn hafa tekið eftir því að krabbameinsfrumur hafa talsvert stytta telómera og að telómerasi er virkur í þessum frumum. Athyglisvert er að telómerasinn varð aðeins virkur eftir að telómerarnir höfðu styst í krabbameinsfrumunum. Ef hægt er að hamla virkni telómerasa í þessum frumum með lyfjum við krabbameinsmeðferð gæti hugsanlega verið hægt að stöðva frekari skiptingu krabbameinsfrumnanna.