14.2 Bygging og raðgreining DNA
14.2 Bygging og raðgreining DNA
Námsmarkmið
Í lok þessa hluta munt þú geta:
- Lýst byggingu DNA
- Útskýrt Sanger-aðferðina við raðgreiningu DNA
- Rætt líkindi og mun á DNA heilkjörnunga og dreifkjörnunga
Byggingareiningar DNA eru kirni. Mikilvægir hlutar kirnis eru niturbasi, 5-kolefna sykra (pentósi) og fosfathópur (mynd 14.5). Kirnið er nefnt eftir niturbasanum. Niturbasinn getur verið púrín, til dæmis adenín (A) og gúanín (G), eða pýrimídín, til dæmis cýtósín (C) og týmín (T).
Púrín hafa tvöfalda hringbyggingu þar sem sexliða hringur er samrunninn fimmliða hring. Pýrimídín eru minni; þau hafa eina sexliða hringbyggingu.
Sykran er deoxýríbósi í DNA og ríbósi í RNA. Kolefnisatóm 5-kolefna sykrunnar eru númeruð 1′, 2′, 3′, 4′ og 5′ (1′ er lesið sem „einn príma“). Fosfatið, sem gerir DNA og RNA súrt, tengist 5′-kolefni sykrunnar þegar estertengi myndast milli fosfórsýru og 5′-OH-hópsins (ester er sýra + alkóhól). Í DNA-kirnum er 3′-kolefni deoxýríbósa tengt hýdroxýlhópi (OH). Í RNA-kirnum inniheldur 2′-kolefni ríbósa einnig hýdroxýlhóp. Basinn tengist 1′-kolefni sykrunnar.
Kirni tengjast hvert öðru og mynda fosfódíestertengi. Fosfatleifin sem er tengd 5′-kolefni sykrunnar í einu kirni myndar annað estertengi við hýdroxýlhópinn á 3′-kolefni sykrunnar í næsta kirni og myndar þannig 5′-3′ fosfódíestertengi. Í fjölkirni hefur annar endi keðjunnar laust 5′-fosfat og hinn endinn lausan 3′-OH-hóp. Þeir kallast 5′-endi og 3′-endi keðjunnar. Basi sem er tengdur 5-kolefna sykru er núkleósíð. Kirni er núkleósíð með einum, tveimur eða þremur fosfathópum.
Á sjötta áratug 20. aldar unnu Francis Crick og James Watson saman að því að ákvarða byggingu DNA við háskólann í Cambridge á Englandi. Aðrir vísindamenn, svo sem Linus Pauling og Maurice Wilkins, rannsökuðu einnig þetta svið. Pauling hafði áður uppgötvað annars stigs byggingu próteina með röntgenkristallafræði. Á rannsóknarstofu Wilkins notaði Rosalind Franklin röntgengeislabeygju til að skilja byggingu DNA. Watson og Crick gátu raðað saman púslinu um DNA-sameindina á grundvelli gagna Franklins, meðal annars vegna þess að Crick hafði einnig rannsakað röntgengeislabeygju (mynd 14.6). Árið 1962 fengu James Watson, Francis Crick og Maurice Wilkins Nóbelsverðlaunin í læknisfræði. Franklin var þá því miður látin, og Nóbelsverðlaun eru ekki veitt eftir andlát.

Watson og Crick lögðu til að DNA væri gert úr tveimur þráðum sem væru snúnir hvor um annan og mynduðu hægri snúinn gorm. Basapörun á sér stað milli púríns og pýrimídíns á gagnstæðum þráðum, þannig að A parast við T og G parast við C, eins og reglur Chargaffs benda til. Þannig eru adenín og týmín samstæð basapör og cýtósín og gúanín eru einnig samstæð basapör. Basapörin eru stöðguð með vetnistengjum: adenín og týmín mynda tvö vetnistengi og cýtósín og gúanín mynda þrjú vetnistengi. Þræðirnir tveir eru andsamsíða; það er, 3′-endi annars þráðarins snýr að 5′-enda hins þráðarins. Sykra og fosfat kirnanna mynda burðargrind byggingarinnar, en niturbasarnir staflast inn á við eins og þrep í stiga. Hvert basapar er aðskilið frá næsta basapari um 0,34 nm og hver snúningur gormsins mælist 3,4 nm. Því eru 10 basapör í hverjum snúningi gormsins. Þvermál tvöfalda DNA-gormsins er 2 nm og er einsleitt eftir allri sameindinni. Aðeins pörun púríns við pýrimídín og andsamsíða stefna DNA-þráðanna tveggja skýra þetta einsleita þvermál. Snúningur þráðanna tveggja hvor um annan myndar jafnt dreifðar stórar og litlar grópir (mynd 14.7).

Raðgreiningartækni fyrir DNA
Fram á tíunda áratug 20. aldar var raðgreining DNA, það er lestur á DNA-röð, tiltölulega dýrt og tímafrekt ferli. Notkun geislamerktra kirna jók vandann vegna öryggissjónarmiða. Með þeirri tækni og þeim sjálfvirku vélum sem nú eru tiltækar er ferlið ódýrara og öruggara og hægt er að ljúka því á nokkrum klukkustundum. Fred Sanger þróaði raðgreiningaraðferðina sem var notuð í verkefninu um raðgreiningu erfðamengis mannsins og er enn mikið notuð í dag (mynd 14.8).
Raðgreiningaraðferðin er þekkt sem dídeoxý-keðjustöðvunaraðferðin. Aðferðin byggir á notkun keðjustöðva, dídeoxýkirna (ddNTP). ddNTP eru frábrugðin deoxýkirnum að því leyti að þau vantar lausan 3′-OH-hóp á 5-kolefna sykrunni. Ef ddNTP er bætt við vaxandi DNA-þráð getur keðjan ekki lengst frekar, því lausi 3′-OH-hópurinn sem þarf til að bæta við öðru kirni er ekki til staðar. Með fyrirfram ákveðnu hlutfalli deoxýkirna og dídeoxýkirna er hægt að mynda DNA-búta af mismunandi stærð.
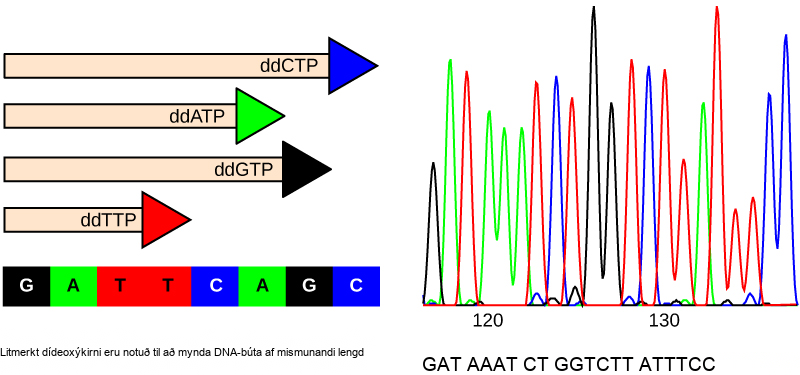
DNA-sýnið sem á að raðgreina er eðlissvipt, það er aðskilið í tvo þræði með hitun við hátt hitastig. DNA er skipt í fjögur glös og í þau er bætt vísi, DNA-pólýmerasa og öllum fjórum núkleósíðþrífosfötunum (A, T, G og C). Að auki er takmörkuðu magni af einu af fjórum dídeoxýnúkleósíðþrífosfötum (ddCTP, ddATP, ddGTP og ddTTP) bætt í hvert glas. Glösin eru merkt A, T, G og C eftir því hvaða ddNTP var bætt við. Til greiningar ber hvert dídeoxýkirnanna fjögurra mismunandi flúrljómandi merki. Keðjulenging heldur áfram þar til flúrljómandi dídeoxýkirni er fellt inn; eftir það verður engin frekari lenging. Þegar hvarfinu er lokið er rafdráttur framkvæmdur. Jafnvel eins basa lengdarmunur er greinanlegur. Röðin er lesin með leysiskanna sem nemur flúrljómandi merki hvers búts. Fyrir vinnu sína við DNA-raðgreiningu fékk Sanger Nóbelsverðlaunin í efnafræði árið 1980.
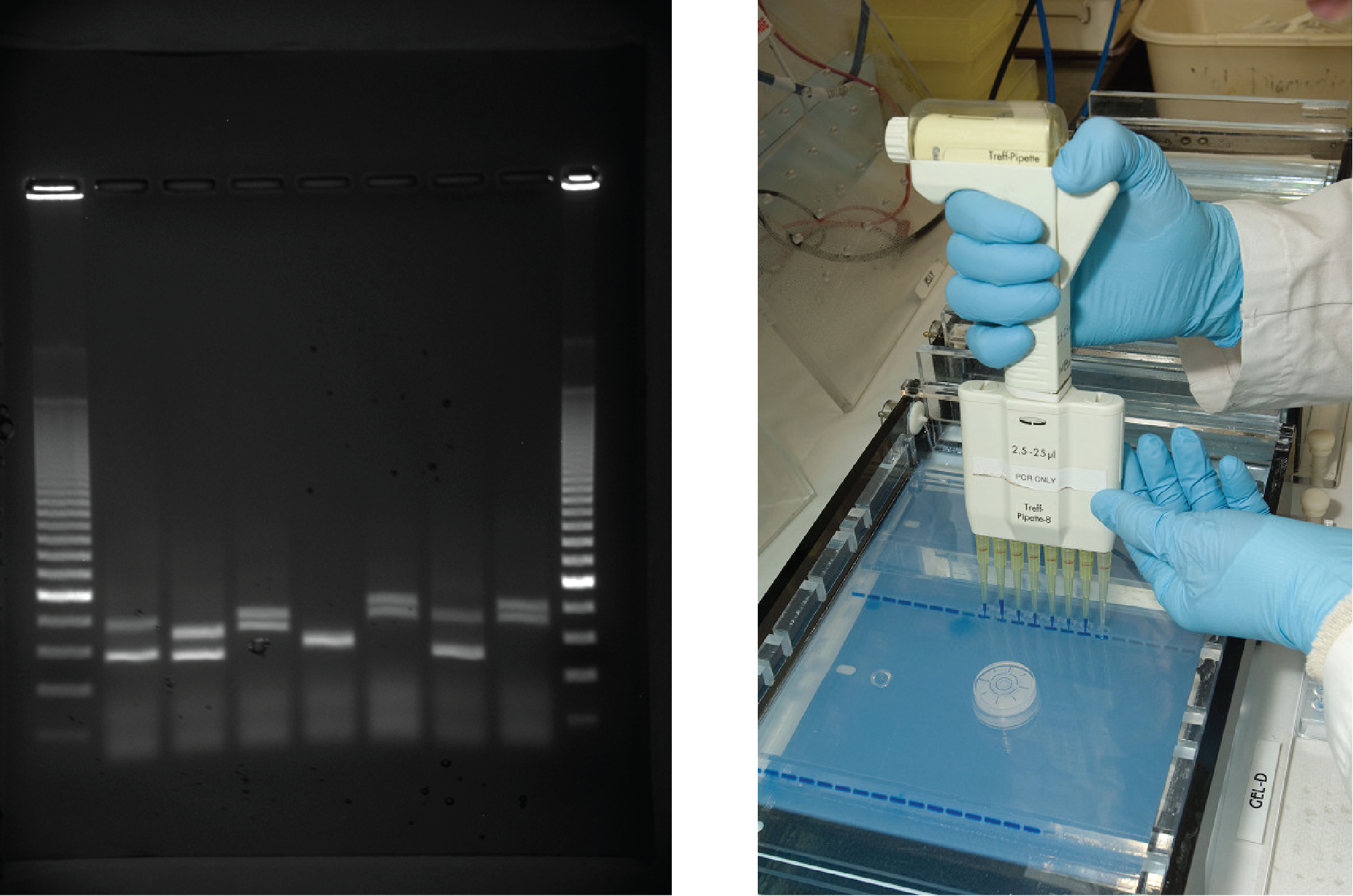
Rafdráttur í geli er tækni sem notuð er til að aðskilja DNA-búta af mismunandi stærð. Yfirleitt er gelið gert úr efni sem kallast agarósi, fjölsykrufjölliðu sem er unnin úr þörungum og er rík af galaktósaleifum. Agarósadufti er bætt í jafnalausn og það hitað. Eftir kælingu er gellausninni hellt í mót. Þegar gelið hefur storknað er DNA sett í gelið og rafstraumi hleypt á. DNA hefur nettóneikvæða hleðslu og færist frá neikvæða rafskautinu í átt að því jákvæða. Rafstraumnum er beitt nógu lengi til að DNA aðskiljist eftir stærð; minnstu bútarnir fara lengst frá brunninum þar sem DNA var sett í gelið, en þyngri bútar með meiri mólmassa haldast næst brunninum. Þegar DNA hefur aðskilist er gelið litað með DNA-sérhæfðu litarefni til skoðunar (mynd 14.9).
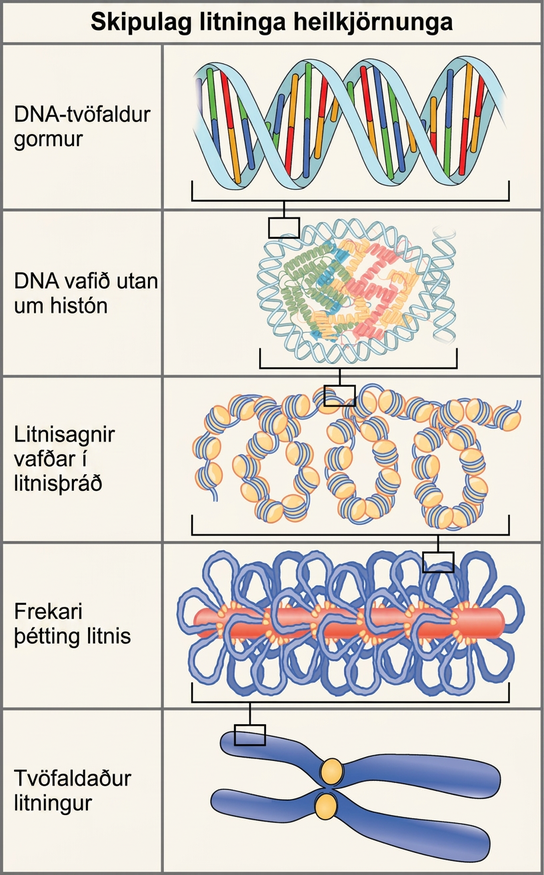
Pökkun DNA í frumum
Dreifkjörnungar eru mun einfaldari en heilkjörnungar að mörgu leyti (mynd 14.10). Flestir dreifkjörnungar innihalda einn hringlaga litning sem er á svæði í umfryminu sem kallast kjarnasvæði.
Stærð erfðamengis eins best rannsakaða dreifkjörnungsins, E. coli, er 4,6 milljónir basapara, eða um það bil 1,1 mm ef það væri klippt og teygt út. Hvernig kemst það þá fyrir í lítilli bakteríufrumu? DNA er snúið með ferli sem kallast ofurvinding. Ofurvinding merkir að DNA er annaðhvort vansnúið (færri en einn snúningur gormsins á hver 10 basapör) eða ofsnúið (fleiri en einn snúningur á hver 10 basapör) miðað við eðlilegt slakað ástand. Vitað er að sum prótein taka þátt í ofurvindingu; önnur prótein og ensím, svo sem DNA-gýrasi, hjálpa til við að viðhalda ofursnúinni byggingu.
Heilkjörnungar, þar sem hver litningur samanstendur af línulegri DNA-sameind, nota annars konar pökkunaraðferð til að koma DNA sínu fyrir inni í kjarnanum (mynd 14.11). Á grunnstigi er DNA vafið utan um prótein sem kallast histón og myndar byggingar sem kallast litnisagnir. Histón eru prótein sem hafa varðveist í þróun, eru rík af basískum amínósýrum og mynda áttliðu úr tveimur sameindum af hverju fjögurra mismunandi históna. Samsetning þeirra og eiginleikar eru mikilvæg til að skilja tjáningu gena og komu að hluta í ljós með rannsóknum Marie M. Daly og Alfred E. Mirsky snemma á sjötta áratug 20. aldar. DNA, sem er neikvætt hlaðið vegna fosfathópanna, er vafið þétt utan um histónkjarnann. Þessi litnisögn tengist þeirri næstu með hjálp tengi-DNA. Þetta kallast einnig „perlur á bandi“-bygging. Með hjálp fimmta histónsins er strengur litnisagna þjappað frekar saman í 30 nm þráð, sem er þvermál byggingarinnar. Metafasalitningar þéttast enn frekar með tengslum við grindarprótein. Á metafasa eru litningarnir þéttastir, um það bil 700 nm á breidd.
Í millifasa hafa litningar heilkjörnunga tvö aðskilin svæði sem hægt er að greina með litun. Þéttpakkaða svæðið kallast þéttlitni og minna þétta svæðið kallast dreiflitni. Þéttlitni inniheldur yfirleitt gen sem eru ekki tjáð og finnst á svæðum þráðhaftsins og litningsendanna. Dreiflitni inniheldur yfirleitt gen sem eru umrituð; þar er DNA pakkað utan um litnisagnir en ekki þjappað frekar saman.