15.5 Ríbósóm og próteinmyndun
15.5 Ríbósóm og próteinmyndun
Markmið náms
Í lok þessa hluta muntu geta gert eftirfarandi:
Próteinmyndun notar meiri orku frumunnar en nokkurt annað efnaskiptaferli. Á móti mynda prótín meiri massa en nokkur annar hluti lífvera, að vatni undanskildu, og prótín sinna nánast öllum hlutverkum frumunnar. Ferlið þýðing, eða próteinmyndun, felur í sér afkóðun mRNA-boða yfir í fjölpeptíðafurð. Amínósýrur eru tengdar saman með samgildum peptíðtengjum í keðjum sem eru frá um það bil 50 upp í meira en 1.000 amínósýruleifar. Hver einstök amínósýra hefur amínóhóp (NH2) og karboxýlhóp (COOH). Fjölpeptíð myndast þegar amínóhópur einnar amínósýru myndar amíðtengi, það er peptíðtengi, við karboxýlhóp annarrar amínósýru (Mynd 15.15). Þetta hvarf er hvatað af ríbósómum og myndar eina vatnssameind.

Vélbúnaður próteinmyndunar
Auk mRNA-mótsins leggja margar sameindir og stórsameindir sitt af mörkum til þýðingarferlisins. Samsetning hvers þáttar getur verið breytileg milli tegunda; til dæmis geta ríbósóm samanstaðið af mismörgum rRNA-sameindum og fjölpeptíðum eftir lífveru. Almenn bygging og hlutverk próteinmyndunarvélarinnar eru þó sambærileg frá bakteríum til frumna manna. Þýðing krefst mRNA-móts, ríbósóma, tRNA og ýmissa ensímþátta. Athugið að líta má á ríbósóm sem ensím þar sem amínósýrubindistaðirnir eru ákvarðaðir af mRNA.
Tengill í námsefni
Smelltu í gegnum skrefin í þessu gagnvirka PBS-efni til að sjá próteinmyndun í verki.
Ríbósóm
Jafnvel áður en mRNA er þýtt þarf fruman að verja orku í að byggja hvert ríbósóm. Í E. coli eru á hverjum tíma 10.000 til 70.000 ríbósóm í hverri frumu. Ríbósóm er flókin stórsameind sem er samsett úr byggingarlegum og hvatandi rRNA-sameindum og mörgum ólíkum fjölpeptíðum. Í heilkjörnungum er kjarnakornið algerlega sérhæft fyrir myndun og samsetningu rRNA.
Ríbósóm finnast í umfrymi dreifkjörnunga og í umfrymi og á hrjúfu frymisneti heilkjörnunga. Hvatberar og grænukorn hafa einnig eigin ríbósóm í hvatberamerg og grænukornagrunni, sem líkjast frekar ríbósómum dreifkjörnunga, og hafa svipað næmi fyrir lyfjum, en ríbósómunum rétt utan við ytri himnur þeirra í umfryminu. Ríbósóm klofna í stóra og litla undireiningu þegar þau eru ekki að mynda prótín og tengjast aftur saman við upphaf þýðingar. Í E. coli er litla undireiningin kölluð 30S og stóra undireiningin 50S, samtals 70S; munið að Svedberg-einingar leggjast ekki saman. Ríbósóm spendýra hafa litla 40S undireiningu og stóra 60S undireiningu, samtals 80S. Litla undireiningin sér um að binda mRNA-mótið en stóra undireiningin bindur tRNA í röð. Hver mRNA-sameind er samtímis þýdd af mörgum ríbósómum sem öll mynda prótín í sömu stefnu: þau lesa mRNA frá 5′ til 3′ og mynda fjölpeptíð frá N-enda til C-enda. Heildarbyggingin úr mRNA og mörgum ríbósómum kallast fjölríbósóm, eða pólýsóm.
tRNA
tRNA eru byggingarlegar RNA-sameindir sem eru umritaðar úr genum af RNA-pólýmerasa III. Eftir tegundum eru 40 til 60 gerðir tRNA til staðar í umfryminu. Tilfærslu-RNA þjóna sem millistykki. Hvert tRNA ber tiltekna amínósýru og þekkir einn eða fleiri mRNA-tákna sem ákvarða röð amínósýra í prótíni. Amínóasýl-tRNA bindast ríbósóminu og bæta samsvarandi amínósýru við fjölpeptíðkeðjuna. tRNA eru því sameindirnar sem í raun „þýða“ tungumál RNA yfir í tungumál prótína.
Af 64 mögulegum mRNA-táknum, það er þríkirnasamsetningum A, U, G og C, tilgreina þrír lok próteinmyndunar og 61 tilgreina viðbót amínósýra við fjölpeptíðkeðjuna. Af þessum 61 tákna kóðar einn tákni (AUG) einnig upphaf þýðingar. Hvert tRNA-andtákni getur basaparast við einn eða fleiri mRNA-tákna fyrir sína amínósýru. Til dæmis myndi röðin CUA á mRNA-móti í réttum lesramma bindast leucín-tRNA sem ber samstæðu röðina GAU. Geta sumra tRNA til að passa við fleiri en einn tákna gefur erfðakóðanum blokkaskipaða byggingu.
Þar sem tRNA eru millistykki þýðingar er merkilegt að þau rúmi svo mikla sértækni í svo litlum pakka. Hafið í huga að tRNA þurfa að víxlverka við þrjá þætti: 1) réttur amínóasýl-tRNA-syntetasi þarf að þekkja þau; 2) ríbósóm þurfa að þekkja þau; og 3) þau þurfa að bindast réttri röð í mRNA.
Amínóasýl-tRNA-syntetasar
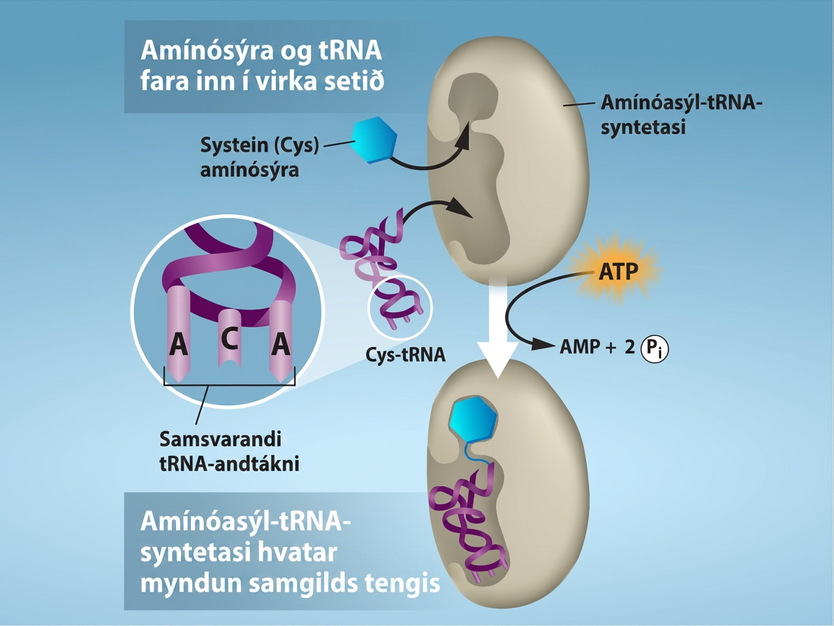
Myndun for-tRNA með RNA-pólýmerasa III býr aðeins til RNA-hluta millistykkjasameindarinnar. Samsvarandi amínósýru þarf að bæta við síðar, þegar tRNA hefur verið unnið og flutt út í umfrymið. Í ferli sem kallast „hleðsla“ tRNA tengir einn úr hópi ensíma, sem kallast amínóasýl-tRNA-syntetasar, hverja tRNA-sameind við rétta amínósýru. Að minnsta kosti ein gerð amínóasýl-tRNA-syntetasa er til fyrir hverja af 20 amínósýrunum; nákvæmur fjöldi þeirra er breytilegur eftir tegund. Þessi ensím binda fyrst og vatnsrjúfa ATP til að hvata myndun orkumikils tengis milli amínósýru og adenósíneinfosfats (AMP); pýrófosfatsameind er rekin út í þessu hvarfi. Virkjaða amínósýran er síðan flutt yfir á tRNA og AMP losnar. Hugtakið „hleðsla“ á vel við, því orkuríka tengið sem festir amínósýru við tRNA er síðar notað til að knýja myndun peptíðtengis. Hvert tRNA er nefnt eftir sinni amínósýru.
Gangverk próteinmyndunar
Eins og mRNA-myndun má skipta próteinmyndun í þrjú stig: upphaf, lengingu og lúkningu. Þýðingarferlið er svipað í dreifkjörnungum og heilkjörnungum. Hér skoðum við hvernig þýðing fer fram í E. coli, dæmigerðum dreifkjörnungi, og tilgreinum mun milli þýðingar í dreifkjörnungum og heilkjörnungum.
Upphaf þýðingar
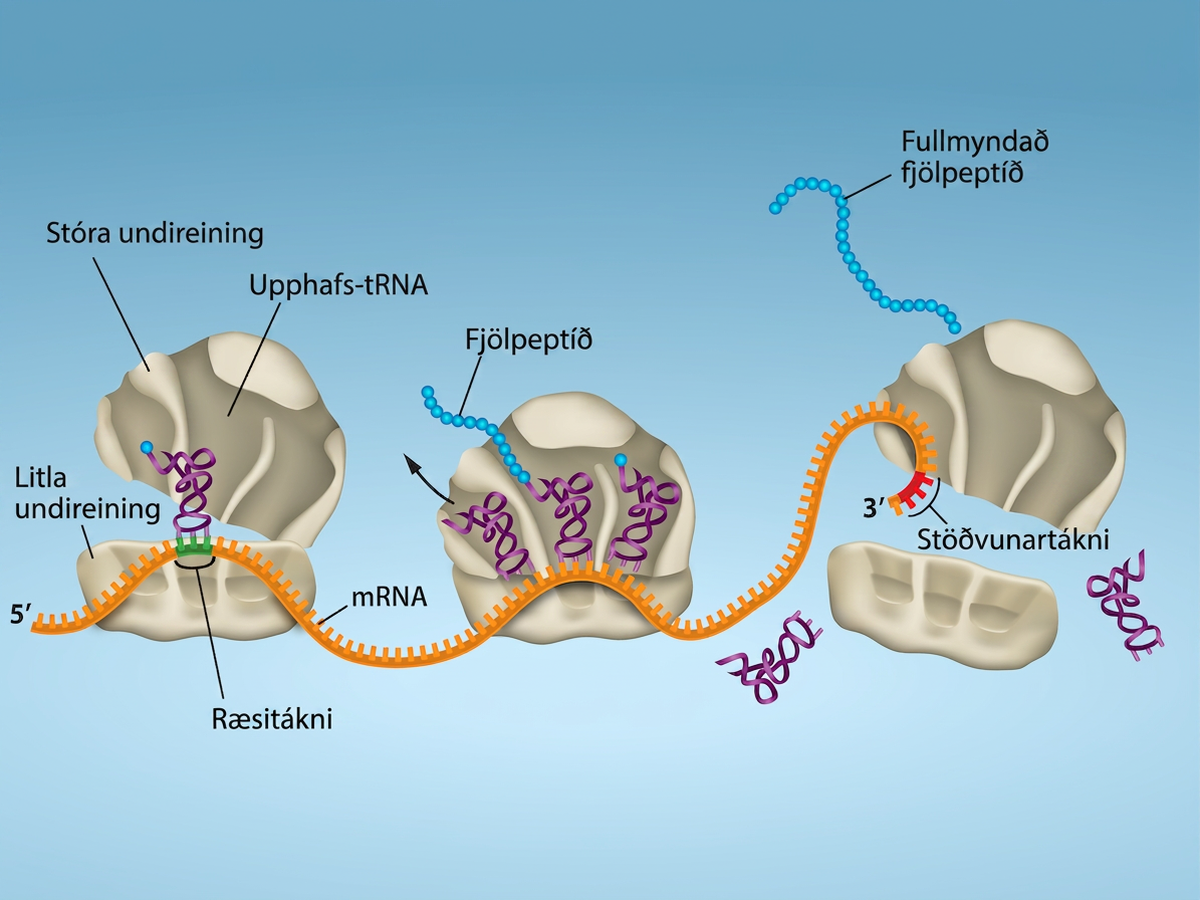
Próteinmyndun hefst með myndun upphafsflóka. Í E. coli felur þessi flóki í sér litla 30S ríbósómið, mRNA-mótið, þrjá upphafsþætti (IF; IF-1, IF-2 og IF-3) og sérstakt upphafs-tRNA sem kallast tRNAᶠᴹᵉᵗ.
Í mRNA E. coli víxlverkar röð ofan við fyrsta AUG-táknann, kölluð Shine-Dalgarno-röð (AGGAGG), við rRNA-sameindirnar sem mynda ríbósómið. Þessi víxlverkun festir 30S ríbósóm-undireininguna á réttum stað á mRNA-mótinu. Gúanósínþrífosfat (GTP), sem er púrínkirnaþrífosfat, þjónar sem orkugjafi við þýðingu, bæði við upphaf lengingar og við tilfærslu ríbósómsins. Binding mRNA við 30S ríbósómið krefst einnig IF-3.
Upphafs-tRNA víxlverkar síðan við ræsitáknann AUG, eða sjaldnar GUG. Þetta tRNA ber amínósýruna metíónín, sem er formýleruð eftir að hún hefur tengst tRNA. Formýleringin myndar gervipeptíðtengi milli formýlkarboxýlhópsins og amínóhóps metíóníns. Binding fMet-tRNAᶠᴹᵉᵗ er miðluð af upphafsþættinum IF-2. fMet byrjar hverja fjölpeptíðkeðju sem E. coli myndar, en er yfirleitt fjarlægt eftir að þýðingu lýkur. Þegar AUG í lesramma kemur fyrir við lengingu þýðingar er óformýlerað metíónín sett inn með venjulegu Met-tRNAᴹᵉᵗ. Eftir myndun upphafsflókans tengist 50S undireiningin við 30S ríbósóm-undireininguna og myndar þýðingarflókann. Í heilkjörnungum myndast svipaður upphafsflóki úr mRNA, litlu 40S ríbósóm-undireiningunni, upphafsþáttum heilkjörnunga og núkleósíðþrífosfötum (GTP og ATP). Metíónínið á hlaðna upphafs-tRNA, sem kallast Met-tRNAᵢ, er ekki formýlerað. Met-tRNAᵢ er þó frábrugðið öðrum Met-tRNA að því leyti að það getur bundið upphafsþætti.
Í stað þess að setjast á Shine-Dalgarno-röðina þekkir upphafsflóki heilkjörnunga 7-metýlgúanósínhettuna á 5′-enda mRNA. Hettubindiprótein (CBP) og nokkrir aðrir upphafsþættir hjálpa ríbósóminu að færast að 5′-hettunni. Þegar upphafsflókinn er kominn að hettunni færist hann eftir mRNA í 5′ til 3′ stefnu og leitar að AUG-ræsitáknanum. Mörg mRNA heilkjörnunga eru þýdd frá fyrsta AUG, en það er ekki alltaf raunin. Samkvæmt Kozak-reglum sýna kirnin í kringum AUG hvort það sé rétti ræsitákninn. Kozak-reglur segja að eftirfarandi samkomulagsröð þurfi að vera í kringum AUG í genum hryggdýra: 5′-gccRccAUGG-3′. R, fyrir púrín, táknar stað sem getur verið annaðhvort A eða G en ekki C eða U. Í meginatriðum er skilvirkni þýðingar meiri eftir því sem röðin er nær þessari samkomulagsröð.
Þegar viðeigandi AUG hefur fundist losna hin prótínin og CBP frá, og 60S undireiningin binst flóka Met-tRNAᵢ, mRNA og 40S undireiningarinnar. Þetta skref lýkur upphafi þýðingar í heilkjörnungum.
Þýðing, lenging og lúkning
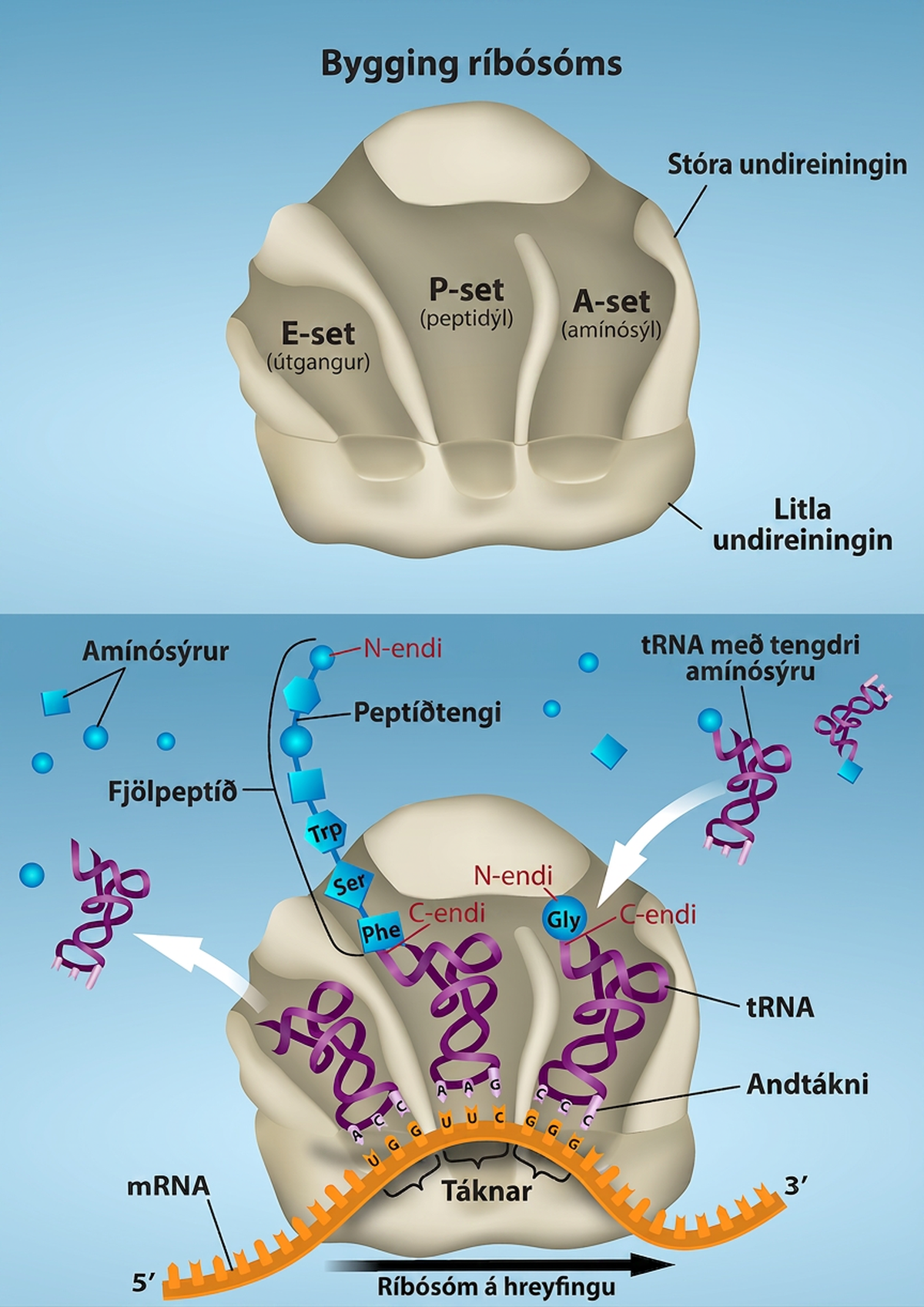
Í dreifkjörnungum og heilkjörnungum eru grunnatriði lengingar þau sömu, svo hér verður lenging skoðuð frá sjónarhóli E. coli. Þegar þýðingarflókinn hefur myndast samanstendur tRNA-bindisvæði ríbósómsins af þremur hólfum. A-setið, eða amínóasýlsetið, bindur komandi hlaðin amínóasýl-tRNA. P-setið, eða peptidýlsetið, bindur hlaðin tRNA sem bera amínósýrur sem hafa myndað peptíðtengi við vaxandi fjölpeptíðkeðjuna en hafa ekki enn losnað frá samsvarandi tRNA. E-setið, eða útgöngusetið, sleppir losnuðum tRNA svo hægt sé að hlaða þau aftur með frjálsum amínósýrum. Upphaflega metíónýl-tRNA situr þó á P-setinu í upphafi lengingarfasa þýðingar bæði í dreifkjörnungum og heilkjörnungum.
Við lengingu þýðingar veitir mRNA-mótið sértækni fyrir tRNA-bindingu. Þegar ríbósómið færist eftir mRNA kemur hver mRNA-tákni í rétta stöðu og tryggt er sértækt bindisamband við samsvarandi hlaðið tRNA-andtákni. Ef mRNA væri ekki til staðar í lengingarflókanum myndi ríbósómið binda tRNA ósértækt og tilviljanakennt.
Lenging heldur áfram með því að hlaðin tRNA fara inn í og yfirgefa ríbósómið í röð eftir því sem hver ný amínósýra er bætt við fjölpeptíðkeðjuna. Færsla tRNA frá A-seti til P-setis og síðan til E-setis verður vegna byggingarbreytinga sem færa ríbósómið áfram um þrjá basa í 3′ stefnu. Orkan fyrir hvert skref eftir ríbósóminu kemur frá lengingarþáttum sem vatnsrjúfa GTP. GTP-orka er nauðsynleg bæði fyrir bindingu nýs amínóasýl-tRNA við A-setið og fyrir tilfærslu þess til P-setisins eftir myndun peptíðtengis. Peptíðtengi myndast milli amínóhóps amínósýrunnar sem tengd er tRNA á A-setinu og karboxýlhóps amínósýrunnar sem tengd er tRNA á P-setinu. Myndun hvers peptíðtengis er hvötuð af peptidýl-transferasa, RNA-byggðu ensími sem er hluti af 50S ríbósóm-undireiningunni. Orkan fyrir myndun hvers peptíðtengis kemur frá orkuríka tenginu sem tengir hverja amínósýru við tRNA hennar. Eftir myndun peptíðtengis færist tRNA á A-setinu, sem nú heldur á vaxandi peptíðkeðjunni, yfir á P-setið, og tRNA á P-setinu, sem nú er tómt, færist yfir á E-setið og er ýtt út úr ríbósóminu (Mynd 15.18). Ótrúlegt er að þýðingarbúnaður E. coli tekur aðeins 0,05 sekúndur í að bæta við hverri amínósýru, sem þýðir að 200 amínósýra prótín má þýða á aðeins 10 sekúndum.
Sjónræn tengsl
Mörg sýklalyf hamla próteinmyndun baktería. Til dæmis lokar tetracýklín A-setinu á bakteríuríbósóminu og klóramfenikól hindrar peptidýl-flutning. Hvaða sértæku áhrif myndir þú búast við að hvort sýklalyf hefði á próteinmyndun?
Tetracýklín hefði bein áhrif á:
Klóramfenikól hefði bein áhrif á:
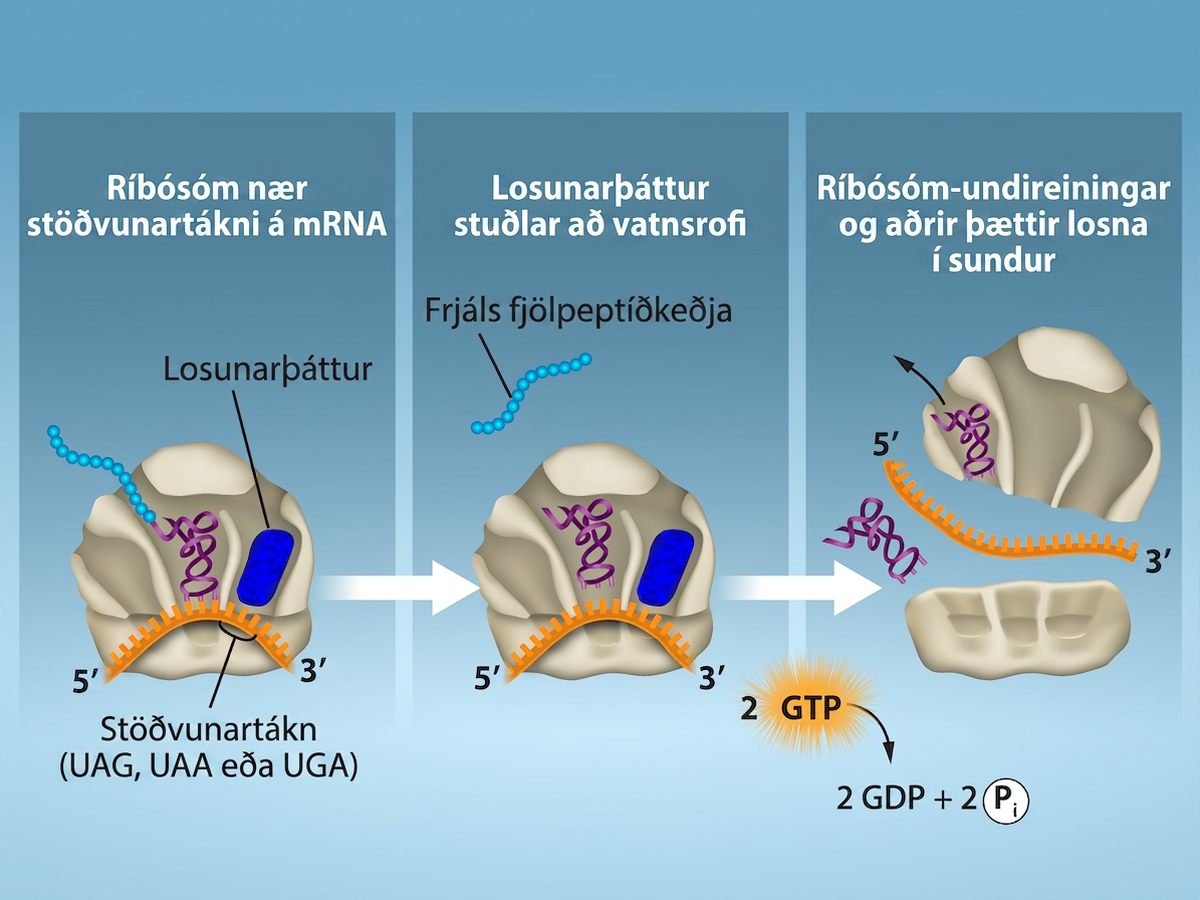
Lúkning þýðingar á sér stað þegar komið er að markleysutákna (UAA, UAG eða UGA). Þegar þessir markleysutáknar raðast upp við A-setið þekkjast þeir af próteinlosunarþáttum sem líkjast tRNA. Losunarþættir í bæði dreifkjörnungum og heilkjörnungum gefa peptidýl-transferasa fyrirmæli um að bæta vatnssameind við karboxýlenda amínósýrunnar á P-setinu. Þetta hvarf neyðir amínósýruna á P-setinu til að losna frá tRNA sínu og nýmyndaða prótínið losnar. Litla og stóra ríbósóm-undireiningin losna frá mRNA og hvor frá annarri; þær eru nánast samstundis kallaðar inn í annan upphafsflóka þýðingar. Eftir að mörg ríbósóm hafa lokið þýðingu er mRNA brotið niður svo hægt sé að endurnýta kirnin í öðru umritunarhvarfi.
Felling, breytingar og miðun prótína
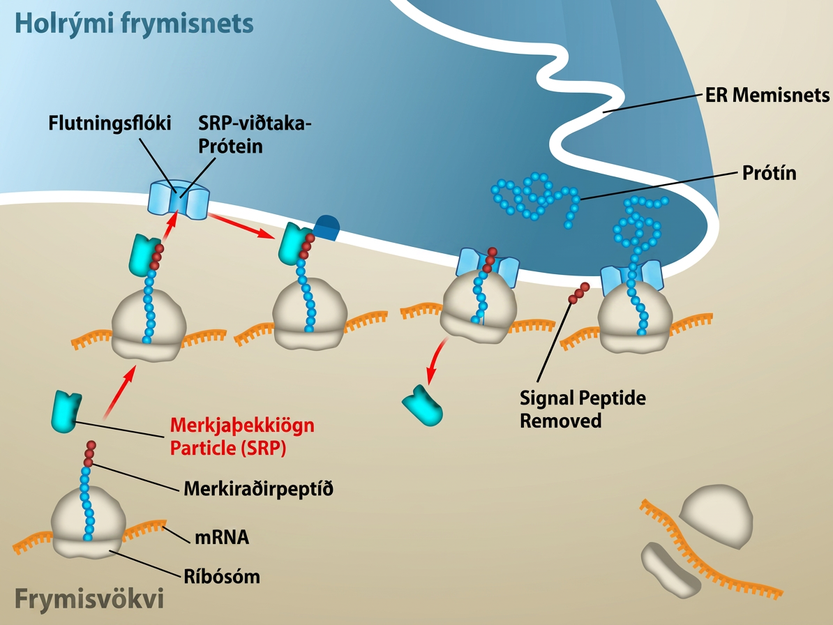
Meðan á þýðingu stendur og eftir hana geta einstakar amínósýrur breyst efnafræðilega, merkiröðum verið bætt við og nýja prótínið „fallið saman“ í sérstaka þrívíða byggingu vegna víxlverkana innan sameindarinnar. Merkiröð er stutt röð við amínóenda prótíns sem beinir því til tiltekins frumuhólfs. Líta má á slíkar raðir sem „lestarmiða“ prótínsins að lokastaðnum, og þær eru þekktar af merkiröðarþekkjandi prótínum sem virka eins og lestarstjórar. Til dæmis mun tiltekinn merkiraðarendi beina prótíni til hvatbera eða grænukorna í plöntum. Þegar prótínið nær áfangastað sínum í frumunni er merkiröðin yfirleitt klippt af.
Mörg prótín falla saman sjálfkrafa, en sum prótín þurfa hjálparsameindir, sem kallast fylgdarprótín, til að koma í veg fyrir að þau kekkist saman í flóknu fellingarferlinu. Jafnvel þótt prótín sé rétt tilgreint af samsvarandi mRNA getur það tekið á sig algjörlega óstarfhæfa lögun ef óeðlilegt hitastig eða pH-skilyrði hindra rétta fellingu.