15.4 Verkun RNA í heilkjörnungum
15.4 RNA-vinnsla í heilkjörnungum
Markmið náms
Í lok þessa hluta muntu geta gert eftirfarandi:
Eftir umritun þurfa for-mRNA heilkjörnunga að ganga í gegnum nokkur vinnsluskref áður en hægt er að þýða þau. tRNA og rRNA í heilkjörnungum, og einnig í dreifkjörnungum, fara líka í gegnum vinnslu áður en þau geta starfað sem hlutar próteinmyndunarvélarinnar.
mRNA-vinnsla
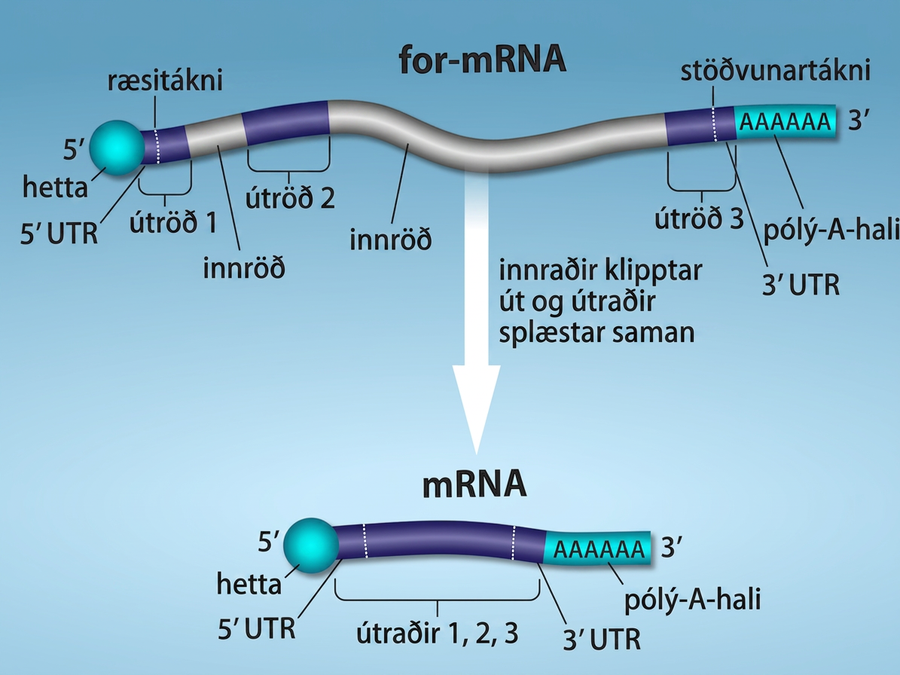
For-mRNA heilkjörnunga fer í gegnum umfangsmikla vinnslu áður en það er tilbúið til þýðingar. Prótínkóðandi raðir heilkjörnunga eru ekki samfelldar eins og í dreifkjörnungum. Kóðandi raðirnar, útraðir, eru rofnar af ókóðandi innröðum sem þarf að fjarlægja til að mynda þýðanlegt mRNA. Viðbótarskrefin í þroskun mRNA heilkjörnunga mynda einnig sameind með mun lengri helmingunartíma en mRNA dreifkjörnunga. mRNA heilkjörnunga endist í nokkrar klukkustundir, en dæmigert mRNA í E. coli endist ekki lengur en fimm sekúndur.
For-mRNA eru fyrst húðuð RNA-stöðugleikaprótínum; þau verja for-mRNA gegn niðurbroti á meðan það er unnið og flutt út úr kjarnanum. Þrjú mikilvægustu skrefin í vinnslu for-mRNA eru viðbót stöðgandi og merkjandi þátta við 5′- og 3′-enda sameindarinnar og fjarlæging innraðanna (Mynd 15.11). Í sjaldgæfum tilvikum er hægt að „ritbreyta“ mRNA-umritinu eftir umritun.
Þróunartengsl
RNA-ritbreyting í trypanosómum
Trypanosómar eru hópur frumdýra sem inniheldur sýkilinn Trypanosoma brucei, sem veldur nagana í nautgripum og svefnsýki í mönnum á stórum svæðum í Afríku (Mynd 15.12). Trypanosóminn berst með bitflugum af ættkvíslinni Glossina, sem oft eru kallaðar tsetse-flugur. Trypanosómar, og nánast allir aðrir heilkjörnungar, hafa frumulíffæri sem kallast hvatberar og sjá frumunni fyrir efnaorku. Hvatberar tjá sitt eigið DNA og eru taldir vera leifar samlífissambands milli heilkjörnungs og innlimaðs dreifkjörnungs. Hvatbera-DNA trypanosóma sýnir áhugaverða undantekningu frá megindogmanu: for-mRNA þeirra hafa ekki réttar upplýsingar til að tilgreina starfhæft prótín. Oftast er ástæðan sú að mRNA vantar nokkur U-kirni. Fruman framkvæmir þá viðbótar RNA-vinnsluskref sem kallast RNA-ritbreyting til að leiðrétta þetta.
Önnur gen í hvatberaerfðamenginu kóða fyrir 40 til 80 kirna leiðar-RNA. Ein eða fleiri þessara sameinda víxlverka með samstæðri basapörun við sum kirni í for-mRNA-umritinu. Leiðar-RNA hefur þó fleiri A-kirni en for-mRNA hefur U-kirni til að bindast við. Á þessum svæðum myndar leiðar-RNA lykkju. 3′-endar leiðar-RNA hafa langan pólý-U-hala og þessir U-basar eru settir inn á þau svæði for-mRNA-umritsins þar sem leiðar-RNA myndar lykkju. Ferlið er alfarið miðlað af RNA-sameindum. Það þýðir að leiðar-RNA, frekar en prótín, virka sem hvatar í RNA-ritbreytingu.
RNA-ritbreyting er ekki aðeins fyrirbæri í trypanosómum. Í hvatberum sumra plantna eru nær öll for-mRNA ritbreytt. RNA-ritbreyting hefur einnig fundist í spendýrum, svo sem rottum, kanínum og jafnvel mönnum. Hver gæti þróunarfræðileg ástæða þessa viðbótarskrefs í vinnslu for-mRNA verið? Einn möguleiki er að hvatberar, sem eru leifar fornra dreifkjörnunga, hafi jafnforna RNA-byggða aðferð til að stjórna tjáningu gena. Til stuðnings þessari tilgátu eru ritbreytingar á for-mRNA ólíkar eftir ástandi frumunnar. Þótt þetta sé tilgáta gæti RNA-ritbreyting verið leif frá frumstæðum tíma þegar RNA-sameindir, fremur en prótín, báru ábyrgð á að hvata efnahvörf.
5′-hettun
Á meðan for-mRNA er enn í myndun er 7-metýlgúanósínhettu bætt við 5′-enda vaxandi umritsins með fosfattengi. Þessi virki hópur ver nýmyndað mRNA gegn niðurbroti. Auk þess þekkja þættir sem taka þátt í próteinmyndun hettuna og hjálpa ríbósómum að hefja þýðingu.
3′ pólý-A-hali
Þegar lengingu er lokið klýfur innkirnaskeri for-mRNA milli AAUAAA-samkomulagsraðar og GU-ríkrar raðar, þannig að AAUAAA-röðin verður eftir á for-mRNA. Ensím sem kallast pólý-A-pólýmerasi bætir síðan við röð af um það bil 200 A-leifum sem kallast pólý-A-hali. Þessi breyting ver for-mRNA enn frekar gegn niðurbroti og er einnig bindistaður fyrir prótín sem er nauðsynlegt til að flytja unnið mRNA út í umfrymið.
Splæsing for-mRNA
Gen heilkjörnunga eru samsett úr útröðum, sem samsvara prótínkóðandi röðum, og innskotsröðum sem kallast innraðir. Innraðir geta tekið þátt í stjórnun gena en eru fjarlægðar úr for-mRNA við vinnslu. Innraðaraðir í mRNA kóða ekki fyrir starfhæfum prótínum.
Uppgötvun innraða kom rannsakendum á óvart á áttunda áratugnum, þar sem þeir bjuggust við að for-mRNA myndu tilgreina prótínraðir án frekari vinnslu, eins og þeir höfðu séð í dreifkjörnungum. Gen æðri heilkjörnunga innihalda mjög oft eina eða fleiri innraðir. Þessi svæði geta samsvarað stýriröðum; þó er líffræðileg þýðing þess að hafa margar innraðir, eða mjög langar innraðir í geni, óljós. Hugsanlegt er að innraðir hægi á tjáningu gena vegna þess að lengri tíma tekur að umrita for-mRNA með mörgum innröðum. Einnig geta innraðir verið óstarfhæfar raðaleifar sem urðu eftir þegar forn gen runnu saman í þróuninni. Þetta er stutt af því að aðskildar útraðir kóða oft fyrir aðskildum undireiningum eða hneppum prótíns. Að mestu leyti geta raðir innraða stökkbreyst án þess að hafa að lokum áhrif á prótínafurðina.
Allar innraðir for-mRNA þarf að fjarlægja algjörlega og nákvæmlega áður en próteinmyndun hefst. Ef ferlið skeikar jafnvel um eitt kirni myndi lesrammi samtengdu útraðanna hliðrast og prótínið sem myndast yrði óstarfhæft. Ferlið við að fjarlægja innraðir og tengja útraðir aftur saman kallast splæsing (Mynd 15.13). Innraðir eru fjarlægðar og brotnar niður á meðan for-mRNA er enn í kjarnanum. Splæsing fer fram með raðsértæku ferli sem tryggir að innraðir séu fjarlægðar og útraðir tengdar saman með nákvæmni upp á eitt kirni. Þótt innröðin sjálf sé ókóðandi eru upphaf og endir hverrar innraðar merkt með tilteknum kirnum: GU við 5′-enda og AG við 3′-enda innraðarinnar. Splæsing for-mRNA er framkvæmd af flókum prótína og RNA-sameinda sem kallast splæsikorn.
Sjónræn tengsl

Villur í splæsingu tengjast krabbameinum og öðrum sjúkdómum í mönnum. Hvers konar stökkbreytingar gætu leitt til splæsingarvillna? Hugsaðu um mismunandi mögulegar afleiðingar ef splæsingarvillur verða.
Athugið að meira en 70 einstakar innraðir geta verið til staðar og hver þeirra þarf að ganga í gegnum splæsingu, auk 5′-hettunar og viðbótar pólý-A-hala, til að mynda eina þýðanlega mRNA-sameind.
Tengill í námsefni
Sjáðu hvernig innraðir eru fjarlægðar við RNA-splæsingu á þessari vefsíðu.
Vinnsla tRNA og rRNA
tRNA og rRNA eru byggingarsameindir sem gegna hlutverkum í próteinmyndun; þessi RNA eru þó ekki sjálf þýdd. For-rRNA eru umrituð, unnin og sett saman í ríbósóm í kjarnakorninu. For-tRNA eru umrituð og unnin í kjarnanum og síðan losuð út í umfrymið, þar sem þau tengjast frjálsum amínósýrum fyrir próteinmyndun.
Flest tRNA og rRNA í heilkjörnungum og dreifkjörnungum eru fyrst umrituð sem löng forverasameind sem nær yfir mörg rRNA eða tRNA. Ensím kljúfa síðan forverana í undireiningar sem samsvara hverju byggingar-RNA. Sumir basar for-rRNA eru metýleraðir, það er -CH3 metýlvirkum hópi er bætt við til stöðugleika. For-tRNA-sameindir ganga einnig í gegnum metýleringu. Eins og hjá for-mRNA fer útskurður undireininga fram í for-RNA heilkjörnunga sem eiga að verða tRNA eða rRNA.
Fullþroska rRNA mynda um það bil 50 prósent af hverju ríbósómi. Sumar RNA-sameindir ríbósómsins eru eingöngu byggingarlegar en aðrar hafa hvata- eða bindivirkni. Fullþroska tRNA taka á sig þrívíða byggingu með staðbundnum svæðum basapörunar sem eru stöðguð með innan-sameinda vetnistengjum. tRNA fellur saman þannig að amínósýrubindistaðurinn er á öðrum endanum og andtáknið á hinum endanum (Mynd 15.14). Andtáknið er þriggja kirna röð í tRNA sem víxlverkar við mRNA-tákna með samstæðri basapörun.
