21.1 Þróun, formgerð og flokkun veira
21.1 Þróun, formgerð og flokkun veira
Námsmarkmið
Í lok þessa hluta munt þú geta gert eftirfarandi:
Veirur eru fjölbreyttar einingar: þær eru ólíkar að byggingu, eftirmyndunaraðferðum og þeim hýslum sem þær sýkja. Næstum öll lífsform hafa veirur sem sýkja þau, allt frá dreifkjörnungum eins og bakteríum og arkeum til heilkjörnunga á borð við plöntur, dýr og sveppi. Þótt hægt sé að skýra mikinn hluta líffræðilegrar fjölbreytni með þróunarsögu, til dæmis hvernig tegundir hafa aðlagast breyttum umhverfisskilyrðum og hvernig tegundir tengjast með sameiginlegum uppruna, er margt enn óþekkt um uppruna og þróun veira.
Uppgötvun og greining
Veirur uppgötvuðust fyrst eftir að postulínssía, Chamberland-Pasteur-sían, var þróuð. Hún gat fjarlægt allar bakteríur sem sáust í smásjá úr vökvasýnum. Árið 1886 sýndi Adolph Meyer fram á að tóbaksmósaíksjúkdómur gæti borist frá veikri tóbaksplöntu til heilbrigðrar með vökvaútdrætti úr plöntu. Árið 1892 sýndi Dmitri Ivanowski að sjúkdómurinn gat borist með þessum hætti jafnvel eftir að Chamberland-Pasteur-sían hafði fjarlægt allar lífvænlegar bakteríur úr útdrættinum. Samt liðu mörg ár áður en sannað var að þessir „síanlegu“ smitvaldar væru ekki einfaldlega mjög smáar bakteríur heldur ný gerð örsmárra sjúkdómsvaldandi agna.
Flestar veiruagnir, eða stakar þroskaðar veirur, eru mjög smáar, um 20 til 250 nanómetrar í þvermál. Nokkrar nýlega uppgötvaðar veirur úr amöbum geta þó orðið allt að 1000 nm í þvermál. Að undanskildum stórum veiruögnum, til dæmis bólusóttarveirum og öðrum stórum DNA-veirum, sjást veirur ekki í ljóssmásjá. Það var ekki fyrr en rafeindasmásjáin var þróuð seint á fjórða áratug 20. aldar að vísindamenn fengu fyrst góða mynd af byggingu tóbaksmósaíkveirunnar (TMV), sem fjallað var um hér að framan (mynd 21.1), og annarra veira (mynd 21.2). Yfirborðsgerð veiruagna má skoða bæði með skönnunarrafeindasmásjá og gegnumlýsingarrafeindasmásjá, en innri bygging veira sést aðeins á myndum úr gegnumlýsingarrafeindasmásjá. Rafeindasmásjártækni og önnur tækni hefur gert kleift að uppgötva margar veirur í öllum gerðum lífvera.

Þróun veira
Þótt líffræðingar viti töluvert um hvernig nútímaveirur stökkbreytast og aðlagast er miklu minna vitað um hvernig veirur urðu upphaflega til. Þegar þróunarsaga flestra lífvera er rannsökuð geta vísindamenn stuðst við steingervingaskrá og sambærilegar sögulegar heimildir. Veirur varðveitast hins vegar ekki sem steingervingar, eftir því sem best er vitað. Rannsakendur verða því að álykta um sögu veira út frá rannsóknum á þróun nútímaveira og með lífefnafræðilegum og erfðafræðilegum upplýsingum.
Flestir fræðimenn eru sammála um að veirur eigi sér ekki einn sameiginlegan forföður og að engin ein tilgáta skýri uppruna allra veira. Nokkrar þróunarsviðsmyndir gætu þó skýrt uppruna veira. Ein þeirra, hrörnunartilgátan, leggur til að veirur hafi þróast úr frjálslífandi frumum eða úr innanfrumu-sníkjudreifkjörnungum. Margt er þó enn óljóst um hvernig slíkt ferli hefði getað átt sér stað. Önnur tilgáta, flóttatilgátan eða framsækin tilgáta, leggur til að veirur hafi orðið til úr RNA- og DNA-sameindum, eða sjálfeftirmyndandi einingum sem líkjast stökkulum eða öðrum hreyfanlegum erfðaþáttum, sem sluppu úr hýsilfrumu og öðluðust getu til að komast inn í aðra frumu. Þriðja tilgátan, frumveirutilgátan, leggur til að veirur hafi verið fyrstu sjálfeftirmyndandi einingarnar, áður en fyrstu frumurnar komu fram. Í öllum tilvikum halda veirur líklega áfram að þróast samhliða frumunum sem þær treysta á sem hýsla.
Eftir því sem tækninni fleygir fram kunna vísindamenn að þróa og betrumbæta fleiri tilgátur um uppruna veira. Nýtt rannsóknarsvið, sameindakerfisfræði veira, reynir einmitt að gera það með samanburði á raðgreindu erfðaefni. Rannsakendur á þessu sviði vonast til að skilja uppruna veira betur í framtíðinni, en slík uppgötvun gæti leitt til framfara í meðferð sjúkdóma sem veirur valda.
Formgerð veira
Veirur eru frumulausar, sem þýðir að þær eru líffræðilegar einingar án frumubyggingar. Þær skortir því flesta hluta frumna, svo sem frumulíffæri, ríbósóm og frumuhimnu. Veiruögn samanstendur af kjarna úr kjarnsýru, ytra próteinhylki eða veiruhylki, og stundum ytri hjúp úr próteinum og fosfólípíðhimnum sem eru upprunnar úr hýsilfrumunni. Veirur geta einnig innihaldið viðbótarprótein, til dæmis ensím, inni í veiruhylkinu eða tengd erfðamengi veirunnar. Augljósasti munurinn á veiruættum er fjölbreytileg formgerð þeirra. Athyglisvert er að flækjustig hýsilsins samsvarar ekki endilega flækjustigi veiruagnarinnar. Í raun finnast sumar flóknustu veiruagnirnar hjá bakteríufögum, veirum sem sýkja einföldustu lifandi lífverurnar, bakteríur.
Formgerð
Veirur eru af mörgum stærðum og gerðum, en þessi einkenni eru stöðug innan hverrar veiruættar. Eins og áður kom fram hafa allar veiruagnir erfðamengi úr kjarnsýru sem er umlukið verndandi veiruhylki. Prótein veiruhylkisins eru skráð í erfðamengi veirunnar og kallast hylkiseiningar. Sum veiruhylki eru einfaldar gormlaga byggingar eða margflötungslegar „kúlur“, en önnur eru mjög flókin að gerð (mynd 21.3).
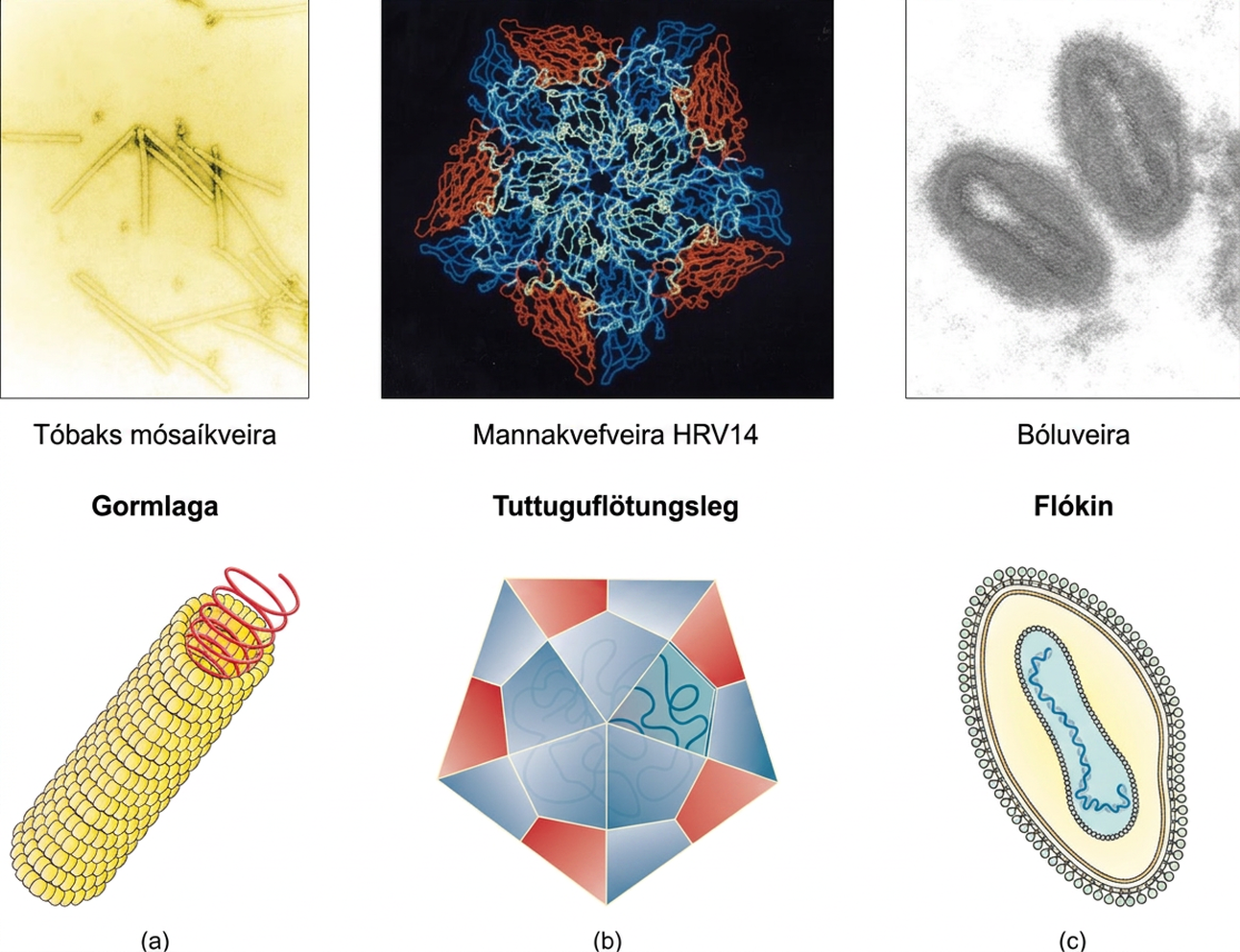
Almennt eru veiruhylki flokkuð í fjóra hópa: gormlaga, tuttuguflötungsleg, hjúpuð og höfuð- og halagerð. Gormlaga veiruhylki eru löng og sívalningslaga. Margar plöntuveirur eru gormlaga, þar á meðal TMV. Tuttuguflötungsveirur eru nærri kúlulaga, til dæmis mænusóttarveira og herpesveirur. Hjúpaðar veirur hafa himnur, upprunnar úr hýsilfrumu, sem umlykja veiruhylkið. Dýraveirur, svo sem HIV, eru oft hjúpaðar. Höfuð- og halaveirur sýkja bakteríur og hafa höfuð sem líkist tuttuguflötungsveirum og hala sem líkist gormlaga veirum.
Margar veirur nota einhvers konar glýkóprótein til að tengjast hýsilfrumum sínum með sameindum á frumuyfirborði sem kallast veiruviðtakar. Hjá þessum veirum er binding nauðsynleg áður en veiran kemst síðar í gegnum frumuhimnuna; aðeins eftir það getur hún lokið eftirmyndun sinni inni í frumunni. Viðtakarnir sem veirur nota eru sameindir sem finnast venjulega á frumuyfirborði og hafa eigin lífeðlisfræðilegt hlutverk. Veirur virðast einfaldlega hafa þróast þannig að þær nýti þessar sameindir fyrir eigin eftirmyndun. HIV notar til dæmis CD4-sameindina á T-eitilfrumum sem einn af viðtökum sínum (mynd 21.4). CD4 er gerð frumuviðloðunarsameindar sem heldur mismunandi tegundum ónæmisfrumna nálægt hver annarri þegar T-eitilfrumuónæmissvar myndast.

Ein flóknasta veiruögn sem þekkt er, T4-bakteríufaginn, sem sýkir bakteríuna Escherichia coli, hefur halabyggingu sem veiran notar til að tengjast hýsilfrumum og höfuðbyggingu sem hýsir DNA hennar.
Adenóveira er óhjúpuð dýraveira sem veldur öndunarfærasjúkdómum í mönnum. Hún notar glýkópróteingadda sem standa út úr hylkiseiningum hennar til að tengjast hýsilfrumum. Óhjúpaðar veirur eru einnig meðal annars þær sem valda mænusótt (mænusóttarveira), ilvörtum (papillomaveira) og lifrarbólgu A (lifrarbólgu A-veira).
Hjúpaðar veiruagnir, til dæmis inflúensuveira, samanstanda af kjarnsýru (RNA í tilfelli inflúensu) og hylkispróteinum sem eru umlukin fosfólípíðtvílagshjúp með próteinum sem veiran skráir fyrir. Glýkóprótein í veiruhjúpnum eru notuð til að tengjast hýsilfrumum. Önnur hjúpprótein eru stoðprótein sem styrkja hjúpinn og gegna oft hlutverki við samsetningu nýrra veiruagna. Hlaupabóla, HIV og hettusótt eru önnur dæmi um sjúkdóma af völdum hjúpaðra veira. Vegna þess hve hjúpurinn er viðkvæmur þola óhjúpaðar veirur betur breytingar á hitastigi, pH-gildi og sum sótthreinsiefni en hjúpaðar veirur.
Í heild segja lögun veiruagnarinnar og tilvist eða fjarvera hjúps lítið um hvaða sjúkdómi veiran getur valdið eða hvaða tegund hún getur sýkt, en þessi einkenni eru samt gagnleg sem upphafspunktur í flokkun veira (mynd 21.5).
Myndatengsl

Hver eftirfarandi fullyrðinga um byggingu veira er sönn?
Rétt er að glýkóprótein hjálpa veirunni að tengjast hýsilfrumunni.
Tegundir kjarnsýra
Ólíkt næstum öllum lifandi lífverum, sem nota DNA sem erfðaefni sitt, geta veirur notað annaðhvort DNA eða RNA. Kjarni veirunnar inniheldur erfðamengið, það er allt erfðaefni veirunnar. Erfðamengi veira eru yfirleitt lítil og innihalda aðeins gen sem skrá fyrir próteinum sem veiran getur ekki fengið frá hýsilfrumunni. Erfðaefnið getur verið einþátta eða tvíþátta. Það getur einnig verið línulegt eða hringlaga. Þótt flestar veirur innihaldi eina kjarnsýru hafa aðrar erfðamengi sem skiptist í nokkra hluta. RNA-erfðamengi inflúensuveirunnar er skipt í hluta. Það stuðlar að breytileika hennar og stöðugri þróun og skýrir hvers vegna erfitt er að þróa bóluefni gegn henni.
Í DNA-veirum beinir veiru-DNA eftirmyndunarpróteinum hýsilfrumunnar til að mynda ný afrit af erfðamengi veirunnar og til að umrita og þýða það erfðamengi yfir í veiruprótein. Meðal sjúkdóma í mönnum af völdum DNA-veira eru hlaupabóla, lifrarbólga B og sjúkdómar af völdum adenóveira. Kynsjúkdómar af völdum DNA-veira eru meðal annars sýkingar af völdum herpesveira og papillomaveiru manna (HPV), sem hefur verið tengd leghálskrabbameini og kynfæravörtum.
RNA-veirur innihalda aðeins RNA sem erfðaefni sitt. Til að eftirmynda erfðamengi sitt í hýsilfrumunni verða RNA-veirur að skrá fyrir eigin ensímum sem geta eftirmyndað RNA yfir í RNA eða, hjá retróveirum, yfir í DNA. Þessi RNA-pólýmerasaensím gera oftar afritunarvillur en DNA-pólýmerasar og gera því oft mistök við umritun. Af þessum sökum verða stökkbreytingar tíðari í RNA-veirum en í DNA-veirum. Það veldur því að þær breytast og aðlagast hýslum sínum hraðar. Meðal sjúkdóma í mönnum af völdum RNA-veira eru inflúensa, lifrarbólga C, mislingar og hundaæði. HIV-veiran, sem smitast kynleiðis, er RNA-retróveira.
Áskorunin við flokkun veira
Þar sem flestar veirur þróuðust líklega frá ólíkum forfeðrum eru kerfisbundnar aðferðir sem vísindamenn hafa notað til að flokka dreifkjörnunga og heilkjörnunga ekki mjög gagnlegar. Ef veirur eru „leifar“ ólíkra lífvera nýtist jafnvel erfðamengis- eða próteingreining takmarkað. Ástæðan er sú að veirur hafa enga sameiginlega erfðamengisröð sem allar deila. Til dæmis er 16S rRNA-röðin, sem er svo gagnleg við að byggja þróunartré dreifkjörnunga, gagnslaus fyrir einingu sem hefur engin ríbósóm. Líffræðingar hafa áður notað nokkur flokkunarkerfi. Í fyrstu voru veirur flokkaðar eftir sameiginlegri formgerð. Síðar voru hópar veira flokkaðir eftir gerð kjarnsýrunnar sem þær innihéldu, DNA eða RNA, og eftir því hvort kjarnsýran var einþátta eða tvíþátta. Þessi eldri flokkunarkerfi röðuðu veirum þó á mismunandi hátt, vegna þess að þau byggðust á ólíkum einkennum veiranna. Algengasta flokkunaraðferðin í dag kallast Baltimore-flokkunarkerfið og byggist á því hvernig boðberi-RNA (mRNA) myndast í hverri veirutegund.
Eldri flokkunarkerfi
Veirur hafa aðeins fá einkenni sem hægt er að nota til flokkunar: erfðamengi veirunnar, gerð veiruhylkis og byggingu hjúpsins hjá hjúpuðum veirum. Öll þessi einkenni hafa áður verið notuð við flokkun veira (tafla 21.1 og mynd 21.6). Erfðamengi veira getur verið breytilegt bæði að gerð erfðaefnis (DNA eða RNA) og skipulagi þess (einþátta eða tvíþátta, línulegt eða hringlaga og skipt eða óskipt). Í sumum veirum eru viðbótarprótein sem þarf til eftirmyndunar tengd beint við erfðamengið eða eru inni í veiruhylkinu.
| Bygging erfðamengis | Dæmi |
|---|---|
| Hundaæðisveira, retróveirur | |
| Herpesveirur, stórubóluveira | |
| Einþátta | Hundaæðisveira, retróveirur |
| Tvíþátta | Herpesveirur, stórubóluveira |
| Línulegt | Hundaæðisveira, retróveirur, herpesveirur, stórubóluveira |
| Hringlaga | Papillomaveirur, margir bakteríufagar |
| Óskipt: erfðamengið samanstendur af einum hluta erfðaefnis | Parainflúensuveirur |
| Skipt: erfðamengið skiptist í marga hluta | Inflúensuveirur |

Veirur má einnig flokka eftir gerð veiruhylkja þeirra (tafla 21.2 og mynd 21.7). Veiruhylki eru flokkuð sem nakin tuttuguflötungsveiruhylki, hjúpuð tuttuguflötungsveiruhylki, hjúpuð gormlaga veiruhylki, nakin gormlaga veiruhylki og flókin veiruhylki. Gerð erfðaefnisins (DNA eða RNA) og bygging þess (einþátta eða tvíþátta, línuleg eða hringlaga og skipt eða óskipt) eru notuð til að flokka byggingu veirukjarnans (tafla 21.2).
| Flokkun veiruhylkja | Dæmi |
|---|---|
| Nakið tuttuguflötungsveiruhylki | Lifrarbólgu A-veira, mænusóttarveirur |
| Hjúpað tuttuguflötungsveiruhylki | Epstein-Barr-veira, herpes simplex-veira, rauðuhundaveira, gulusóttarveira, HIV-1 |
| Hjúpað gormlaga veiruhylki | Inflúensuveirur, hettusóttarveira, mislingaveira, hundaæðisveira |
| Nakið gormlaga veiruhylki | Tóbaksmósaíkveira |
| Flókið veiruhylki með mörgum próteinum; sumar hafa blöndu af tuttuguflötungs- og gormlaga hylkisbyggingum | Herpesveirur, stórubóluveira, lifrarbólgu B-veira, T4-bakteríufagi |

Baltimore-flokkunin
Algengasta og nú mest notaða flokkunarkerfi veira var fyrst þróað af Nóbelsverðlaunalíffræðingnum David Baltimore snemma á áttunda áratug 20. aldar. Auk munar á formgerð og erfðum sem nefndur var hér að ofan flokkar Baltimore-flokkunarkerfið veirur eftir því hvernig mRNA myndast í eftirmyndunarferli veirunnar.
Veirur í hópi I hafa tvíþátta DNA (dsDNA) sem erfðamengi. mRNA þeirra myndast með umritun á mjög svipaðan hátt og hjá frumu-DNA, með ensímum hýsilfrumunnar.
Veirur í hópi II hafa einþátta DNA (ssDNA) sem erfðamengi. Þær breyta einþátta erfðamengi sínu í dsDNA-millistig áður en umritun yfir í mRNA getur átt sér stað.
Veirur í hópi III nota dsRNA sem erfðamengi. Strengirnir aðskiljast og annar þeirra er notaður sem mót fyrir myndun mRNA með RNA-háðum RNA-pólýmerasa sem veiran skráir fyrir.
Veirur í hópi IV hafa ssRNA-erfðamengi með jákvæðum skautunarstreng. Það þýðir að erfðamengis-RNA getur þjónað beint sem mRNA. dsRNA-millistig, sem kallast eftirmyndunarmillistig, verða til þegar erfðamengis-RNA er afritað. Úr þessum millistigum myndast margir heilir RNA-strengir með neikvæðum skautunarstreng, sem eru bætiröð við jákvætt erfðamengis-RNA. Þeir geta síðan þjónað sem mót fyrir myndun RNA með jákvæðum skautunarstreng, bæði heils erfðamengis-RNA og styttri veiru-mRNA-sameinda.
Veirur í hópi V hafa ssRNA-erfðamengi með neikvæðum skautunarstreng, sem þýðir að röð þeirra er bætiröð við mRNA. Eins og hjá veirum í hópi IV eru dsRNA-millistig notuð til að mynda afrit af erfðamenginu og framleiða mRNA. Í þessu tilviki er hægt að breyta neikvæða erfðamengisstrengnum beint í mRNA. Auk þess myndast heilir jákvæðir RNA-strengir sem þjóna sem mót fyrir myndun neikvæða erfðamengisstrengsins.
Veirur í hópi VI hafa tvö eintök af ssRNA-erfðamengi sem verður að breyta í dsDNA með ensíminu öfugum umritara. dsDNA-ið er síðan flutt inn í kjarna hýsilfrumunnar og sett inn í erfðamengi hýsilsins. Þá getur mRNA myndast með umritun á veiru-DNA sem hefur verið innlimað í erfðamengi hýsilsins.
Veirur í hópi VII hafa ófullgerð dsDNA-erfðamengi og mynda ssRNA-millistig sem virka sem mRNA. Þeim er einnig breytt aftur í dsDNA-erfðamengi með öfugum umritara, sem er nauðsynlegt fyrir eftirmyndun erfðamengisins.
Einkenni hvers hóps í Baltimore-flokkuninni eru dregin saman í töflu 21.3 ásamt dæmum um hvern hóp.
| Hópur | Einkenni | Aðferð við myndun mRNA | Dæmi |
|---|---|---|---|
| Tvíþátta DNA (dsDNA) | mRNA er umritað beint af DNA-mótinu | Herpes simplex-veira (herpesveira) | |
| Einþátta DNA (ssDNA) | DNA breytist í tvíþátta form áður en RNA er umritað | Hundaparvóveira (parvóveira) | |
| Tvíþátta RNA (dsRNA) | mRNA er umritað af RNA-erfðamenginu | Iðrakveisa í börnum (rótaveira) | |
| Einþátta RNA (+) | Erfðamengið virkar sem mRNA | Kvef (picornaveira) | |
| V | Einþátta RNA (-) | mRNA er umritað af RNA-erfðamenginu | Hundaæði (rhabdoveira) |
| Einþátta RNA-veirur með öfugan umritara | Öfugur umritari myndar DNA út frá RNA-erfðamenginu; DNA er síðan innlimað í erfðamengi hýsilsins; mRNA er umritað af innlimaða DNA-inu | Eyðniveira manna (HIV) | |
| Tvíþátta DNA-veirur með öfugan umritara | Erfðamengi veirunnar er tvíþátta DNA, en veiru-DNA er eftirmyndað í gegnum RNA-millistig; RNA getur þjónað beint sem mRNA eða sem mót fyrir myndun mRNA | Lifrarbólgu B-veira (hepadnaveira) |