16.5 Stjórnun gena eftir umritun í heilkjörnungum
16.5 Stjórnun gena eftir umritun í heilkjörnungum
Námsmarkmið
Í lok þessa hluta munt þú geta gert eftirfarandi:
RNA er umritað en verður að vinna í fullþroska form áður en þýðing getur hafist. Þessi vinnsla, sem fer fram eftir að RNA-sameind hefur verið umrituð en áður en hún er þýdd í prótín, kallast breyting eftir umritun. Líkt og á stigum utangenaerfða og umritunar má einnig stjórna þessu skrefi eftir umritun til að stýra genatjáningu í frumunni. Ef RNA er ekki unnið, flutt eða þýtt myndast ekkert prótín.
RNA-splæsing, fyrsta stig stjórnunar eftir umritun
Í heilkjörnungafrumum inniheldur RNA-umritið oft svæði, sem kallast innraðir, sem eru fjarlægð fyrir þýðingu. Svæði RNA sem kóða fyrir prótín kallast útraðir (Mynd 16.11). Eftir að RNA-sameind hefur verið umrituð, en áður en hún fer úr kjarnanum til að verða þýdd, er RNA unnið og innraðirnar fjarlægðar með splæsingu. Splæsing fer fram í splæsikornum, ríbókjarnaprótínflókum sem geta þekkt báða enda innraðarinnar, klippt umritið á þessum tveimur stöðum og fært útraðirnar saman til samtengingar.

Tengsl við þróun
Valkvæð RNA-splæsing
Á áttunda áratugnum sáust fyrst gen sem sýndu valkvæða RNA-splæsingu. Valkvæð RNA-splæsing er ferli sem gerir kleift að framleiða mismunandi prótínafurðir úr einu geni þegar mismunandi samsetningar útraða eru sameinaðar til að mynda mRNA (Mynd 16.12). Þessi valkvæða splæsing getur verið tilviljanakennd, en oftar er henni stjórnað og hún virkar sem aðferð við stjórnun gena: fruman stýrir tíðni mismunandi splæsingarvalkosta til að stjórna framleiðslu mismunandi prótínafurða í mismunandi frumum eða á mismunandi þroskastigum. Nú er vitað að valkvæð splæsing er algeng aðferð við stjórnun gena hjá heilkjörnungum; samkvæmt einu mati eru 70 prósent gena í mönnum tjáð sem mörg prótín með valkvæðri splæsingu. Þó að RNA-umritum megi splæsa á marga valkvæða vegu er upprunaleg 5′-3′ röð útraðanna alltaf varðveitt. Umrit með útröðunum 1 2 3 4 5 6 7 gæti því verið splæst sem 1 2 4 5 6 7 eða 1 2 3 6 7, en aldrei sem 1 2 5 4 3 6 7.
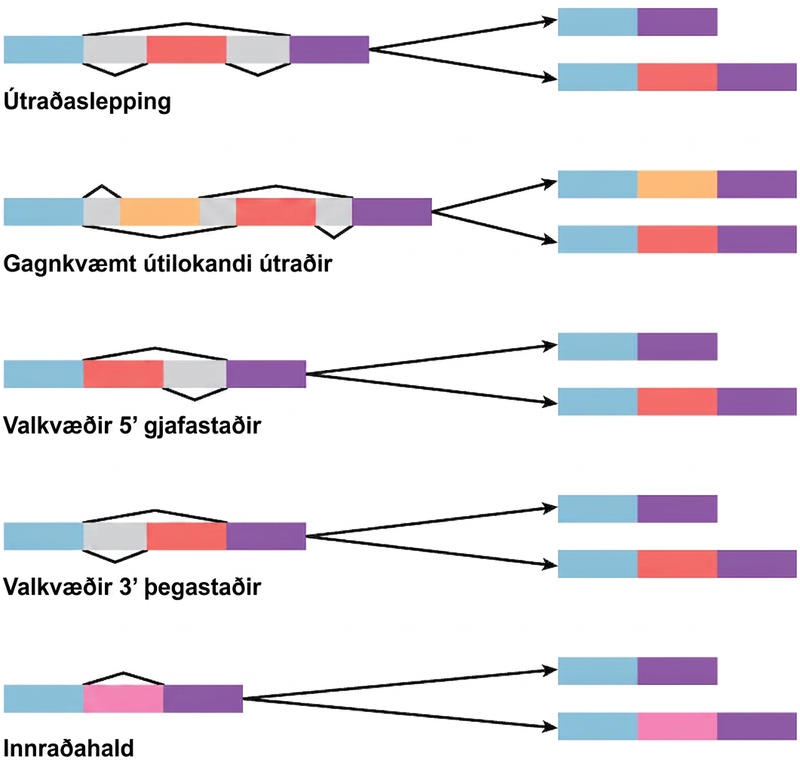
Hvernig gat valkvæð splæsing þróast? Innraðir hafa kenniraðir við upphaf og enda; auðvelt er að ímynda sér að splæsingarkerfið beri ekki kennsl á enda innraðar og finni í staðinn enda næstu innraðar, þannig að tvær innraðir og útröðin á milli þeirra séu fjarlægðar. Reyndar eru til ferli sem koma í veg fyrir slíka útraðasleppingu, en stökkbreytingar geta valdið því að þau bregðist. Slík „mistök“ myndu að öllum líkindum framleiða óvirkt prótín. Orsök margra erfðasjúkdóma er raunar óeðlileg splæsing fremur en stökkbreytingar í kóðunarröð. Valkvæð splæsing gæti hins vegar búið til prótínafbrigði án þess að upprunalega prótínið glatist og þannig opnað möguleika á að nýja afbrigðið aðlagist nýjum hlutverkum. Tvöföldun gena hefur á svipaðan hátt gegnt mikilvægu hlutverki í þróun nýrra hlutverka með því að útvega gen sem getur þróast án þess að upprunalega, virka prótíninu sé útrýmt.
Spurning: Í kornsnáknum Pantherophis guttatus eru nokkur mismunandi litaafbrigði, þar á meðal melanínlausir snákar þar sem húðmynstrið sýnir aðeins rauð og gul litarefni. Orsök melanínleysis í þessum snákum var nýlega rakin til innskots stökkuls inn í innröð í OCA2-geninu, sem tengist augn- og húðalbínisma. Hvernig gæti innskot auka erfðaefnis inn í innröð leitt til óvirks prótíns?
Stjórnun á stöðugleika RNA
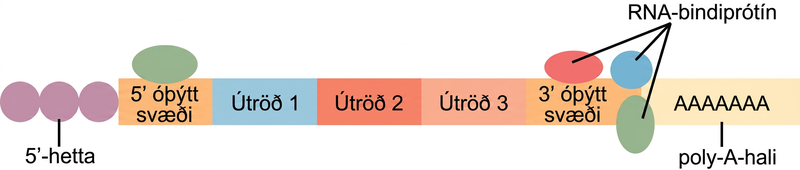
Áður en mRNA yfirgefur kjarnann fær það tvær verndandi „hettur“ sem koma í veg fyrir að endar strengsins brotni niður á ferðalaginu. 5′- og 3′-útkirnaskerar geta brotið niður óvarið RNA. 5′-hettan, sem er sett á 5′-enda mRNA, er venjulega gerð úr metýleraðri gúanósínþrífosfat-sameind (GTP). GTP er sett „afturábak“ á 5′-enda mRNA, þannig að 5′-kolefni GTP og endakirnisins tengjast með þremur fosfötum. Pólý-A-halinn, sem er festur við 3′-endann, er venjulega löng keðja adenínkirna. Þessar breytingar vernda báða enda RNA fyrir árás útkirnaskera.
Þegar RNA hefur verið flutt út í umfrymið má stjórna því hversu lengi það dvelur þar. Hver RNA-sameind hefur skilgreindan líftíma og brotnar niður á tilteknum hraða. Þessi niðurbrotshraði getur haft áhrif á magn prótíns í frumunni. Ef niðurbrotshraðinn eykst er RNA ekki eins lengi til staðar í umfryminu og tíminn sem er tiltækur fyrir þýðingu mRNA styttist. Ef niðurbrotshraðinn minnkar dvelur mRNA-sameindin lengur í umfryminu og meira prótín getur verið þýtt. Þessi niðurbrotshraði er kallaður stöðugleiki RNA. Ef RNA er stöðugt greinist það lengur í umfryminu.
Binding prótína við RNA getur einnig haft áhrif á stöðugleika þess. Prótín sem kallast RNA-bindiprótín, eða RBP, geta bundist svæðum á mRNA rétt ofan eða neðan við prótínkóðandi svæðið. Þessi svæði í RNA, sem eru ekki þýdd í prótín, kallast óþýdd svæði, eða UTR. Þau eru ekki innraðir; þær hafa verið fjarlægðar í kjarnanum. Þess í stað eru þetta svæði sem stjórna staðsetningu mRNA, stöðugleika og próteinþýðingu. Svæðið rétt á undan prótínkóðandi svæðinu kallast 5′ UTR, en svæðið á eftir kóðunarsvæðinu kallast 3′ UTR (Mynd 16.13). Binding RBP við þessi svæði getur aukið eða minnkað stöðugleika RNA-sameindar eftir því hvaða tiltekna RBP binst.
Stöðugleiki RNA og örRNA
Auk RBP, sem bindast RNA og stjórna stöðugleika þess með því að auka hann eða minnka, geta aðrir þættir sem kallast örRNA bundist RNA-sameindinni. Þessi örRNA, eða miRNA, eru stuttar RNA-sameindir, aðeins 21 til 24 kirni að lengd. miRNA verða til í kjarnanum sem lengri for-miRNA. Þessi for-miRNA eru klippt niður í fullþroska miRNA með prótíni sem kallast Dicer. Líkt og umritunarþættir og RBP þekkja fullþroska miRNA tiltekna röð og bindast RNA; miRNA tengjast þó einnig ríbókjarnaprótínflóka sem kallast RNA-hvattur þöggunarflóki (RISC). RNA-hluti RISC basaparast við samsvarandi raðir á mRNA og annaðhvort hindrar þýðingu boðanna eða leiðir til niðurbrots mRNA.