47.1 Kreppa líffræðilegrar fjölbreytni
47.1 Kreppa líffræðilegrar fjölbreytni
Námsmarkmið
Í lok þessa hluta munt þú geta gert eftirfarandi:
Hefðbundið hafa vistfræðingar mælt líffræðilega fjölbreytni, sem er almennt hugtak yfir fjölda tegunda í lífhvolfinu, með því að taka tillit til bæði fjölda tegunda og hlutfallslegrar gnægðar þeirra innbyrðis. Líffræðilega fjölbreytni má meta á nokkrum skipulagsstigum lifandi vera. Þessir matsstuðlar, sem koma úr upplýsingafræði, eru gagnlegastir sem fyrsta skref í að mæla líffræðilega fjölbreytni milli og innan vistkerfa; þeir eru síður gagnlegir þegar helsta áhyggjuefni verndunarlíffræðinga er einfaldlega tap á líffræðilegri fjölbreytni. Hins vegar viðurkenna líffræðingar að mælikvarðar á líffræðilega fjölbreytni, hvað varðar tegundafjölbreytni, geti hjálpað til við að beina aðgerðum að því að vernda þá þætti líffræðilegrar fjölbreytni sem eru mikilvægir líffræðilega eða tæknilega.
Síkliðurnar í Viktoríuvatni gefa dæmi sem hjálpar okkur að skilja líffræðilega fjölbreytni. Líffræðingarnir sem rannsökuðu síkliður á níunda áratug 20. aldar fundu hundruð síkliðutegunda sem höfðu sérhæfst á margvíslegan hátt að ólíkum búsvæðum og fæðuháttum: sumar átu svif sem svífur í vatninu, aðrar skófu eða átu þörunga af steinum, enn aðrar átu skordýralirfur af botni vatnsins og sumar átu egg annarra síkliðutegunda. Síkliðurnar í Viktoríuvatni eru afurð flókinnar aðlögunarkvíslunar. Aðlögunarkvíslun er hröð kvíslun, með tegundamyndun, þar sem þróunarfræðileg ættlína greinist í margar náskyldar tegundir; hjá síkliðum Viktoríuvatns tók þetta innan við þrjár milljónir ára. Tegundirnar „kvíslast“ yfirleitt inn í ólík búsvæði og vistsess. Finkur Galápagos-eyja eru dæmi um hóflega aðlögunarkvíslun með 15 tegundum. Síkliðurnar í Viktoríuvatni eru dæmi um stórbrotna aðlögunarkvíslun sem áður náði til um 500 tegunda.
Á sama tíma og líffræðingar voru að gera þessa uppgötvun tóku sumar tegundir að hverfa hratt. Ein orsökin var nílarkarfinn, stór ránfiskur sem fiskveiðiyfirvöld komu fyrir í Viktoríuvatni til að afla fæðu fyrir fólkið sem bjó umhverfis vatnið. Nílarkarfanum var sleppt í vatnið árið 1963, en stofnar hans tóku ekki að stækka hratt fyrr en á níunda áratugnum. Karfinn stækkaði stofn sinn með því að éta síkliður og rak hverja tegundina á fætur annarri að mörkum útdauða. Reyndar komu nokkrir þættir við sögu í útdauða hugsanlega 200 síkliðutegunda í Viktoríuvatni: nílarkarfinn, minnkandi vatnsgæði vegna landbúnaðar og landhreinsunar við strendur vatnsins og aukinn veiðiþrýstingur. Vísindamenn höfðu ekki einu sinni skráð allar tegundirnar sem voru til staðar; svo margar týndust án þess að hafa nokkurn tíma fengið nafn. Fjölbreytnin er nú aðeins skuggi af því sem hún var áður.
Síkliðurnar í Viktoríuvatni eru smámynd af því hraða tegundatapi sem á sér stað um alla jörð í dag og stafar fyrst og fremst af athöfnum manna. Útdauði er náttúrulegt ferli stórþróunar sem verður að jafnaði á þeim hraða að um ein af hverjum milljón tegundum deyr út á ári. Steingervingasagan sýnir að fimm tímabil fjöldaútdauða hafa orðið í sögunni, með mun meiri tegundatapi, og hraði tegundataps í dag er sambærilegur við slík tímabil. Mikill munur er þó á fyrri fjöldaútdauðum og þeim útdauða sem við upplifum nú: nú ræður athafnasemi manna mestu. Þrjár athafnir manna hafa sérstaklega mikil áhrif: 1) eyðing búsvæða, 2) innleiðing framandi tegunda og 3) ofnýting. Spár um tegundatap á næstu öld, sem er örstuttur tími á jarðsögulegum tímakvarða, eru á bilinu 10 til 50 prósent. Útdauði af þessari stærðargráðu hefur aðeins orðið fimm sinnum áður í sögu jarðar og þeir útdauðar stöfuðu af hamfaraatburðum sem breyttu í hvert sinn framvindu sögu lífsins.
Gerðir líffræðilegrar fjölbreytni
Vísindamenn fallast almennt á að hugtakið líffræðileg fjölbreytni lýsi fjölda og gerðum tegunda og gnægð þeirra á tilteknum stað eða á jörðinni. Erfitt getur verið að skilgreina tegundir, en flestir líffræðingar eru samt sáttir við hugtakið og geta í flestum tilfellum greint og talið heilkjörnungategundir. Líffræðingar hafa einnig skilgreint aðra mælikvarða á líffræðilega fjölbreytni, og sumir þeirra skipta máli þegar skipuleggja á verndun hennar.
Erfðafjölbreytni er einn slíkra mælikvarða. Erfðafjölbreytni, eða erfðabreytileiki, er hráefni þróunar og aðlögunar tegundar. Framtíðarmöguleikar tegundar til aðlögunar ráðast af þeirri erfðafjölbreytni sem er varðveitt í erfðamengjum einstaklinganna í þeim stofnum sem mynda tegundina. Hið sama gildir um hærri flokkunarfræðilega hópa. Ættkvísl með mjög ólíkum tegundum býr yfir meiri erfðafjölbreytni en ættkvísl með erfðafræðilega líkum tegundum sem hafa svipaða vistfræði. Ef velja þyrfti hvorri ættkvíslinni ætti að halda til haga væri sú erfðafræðilega fjölbreyttari líklegri til að hafa meiri möguleika á frekari þróun.
Mörg gen skrá fyrir próteinum, sem aftur sjá um efnaskiptaferla sem halda lífverum á lífi og gera þeim kleift að fjölga sér. Erfðafræðilegan fjölbreytileika má mæla sem efnafræðilegan fjölbreytileika þar sem ólíkar tegundir framleiða fjölbreytt efni í frumum sínum, bæði próteinin sem og afurðir og hliðarafurðir efnaskipta. Þessi efnafræðilegi fjölbreytileiki getur haft mögulegan ávinning fyrir menn sem uppspretta lyfja, svo hann veitir eina leið til að mæla fjölbreytileika sem er mikilvægur fyrir heilsu og velferð manna.
Menn hafa skapað fjölbreytileika í húsdýrum, nytjaplöntum og sveppum, meðal margra annarra lífvera. Þessi fjölbreytileiki verður einnig fyrir tapi vegna fólksflutninga, markaðsafla og aukinnar hnattvæðingar í landbúnaði, sérstaklega á þéttbýlum svæðum eins og í Kína, Indlandi og Japan. Mannkynið er beinlínis háð þessum fjölbreytileika sem stöðugri fæðuuppsprettu, og hnignun hans veldur líffræðingum og landbúnaðarvísindamönnum áhyggjum.
Einnig er gagnlegt að skilgreina vistkerfafjölbreytni, það er fjölda ólíkra vistkerfa á jörðinni eða innan tiltekins landsvæðis (mynd 47.2). Heil vistkerfi geta horfið jafnvel þótt sumar tegundirnar lifi af með því að aðlagast öðrum vistkerfum. Tap vistkerfis þýðir tap á samskiptum tegunda, tap á einstökum einkennum samaðlögunar og tap á líffræðilegri framleiðni sem vistkerfið getur skapað. Dæmi um vistkerfi sem er að mestu horfið í Norður-Ameríku er sléttuvistkerfið. Slétturnar náðu eitt sinn yfir miðhluta Norður-Ameríku frá barrskógunum í norðurhluta Kanada suður til Mexíkó. Nú eru þær nánast horfnar og akrar, beitilönd og úthverfi hafa tekið við. Margar tegundanna lifa annars staðar, en hið gríðarlega framleiðna vistkerfi sem myndaði frjósamasta landbúnaðarjarðveg Bandaríkjanna er horfið. Afleiðingin er sú að upprunalegur jarðvegur er að hverfa eða þarf að viðhalda honum og bæta hann með miklum tilkostnaði.

Þrátt fyrir mikla viðleitni er þekking á þeim tegundum sem byggja jörðina takmörkuð og verður það alltaf vegna viðvarandi skorts á fjármagni og pólitískum vilja. Nýlegt mat bendir til þess að þær heilkjörnungategundir sem vísindin hafa gefið nöfn, um 1,5 milljón tegunda, séu innan við 20 prósent af heildarfjölda heilkjörnungategunda á jörðinni (8,7 milljónir tegunda samkvæmt einu mati). Áætlanir um fjölda dreifkjörnungategunda eru að mestu ágiskanir, en líffræðingar eru sammála um að vísindin séu rétt að byrja að skrá fjölbreytni þeirra. Jafnvel þegar litið er til þess sem vitað er, er engin miðlæg geymsla til fyrir nöfn eða sýni lýstra tegunda; því er engin leið að vera viss um að þessar 1,5 milljón lýsingar séu nákvæm talning. Þetta er besta ágiskun byggð á áliti sérfræðinga í ólíkum flokkunarhópum. Í ljósi þess að jörðin tapar tegundum með vaxandi hraða eru vísindin að miklu leyti í sömu stöðu og þau voru gagnvart síkliðunum í Viktoríuvatni: þau vita lítið um það sem er að tapast. Tafla 47.1 sýnir nýlegar áætlanir um líffræðilega fjölbreytni í mismunandi hópum.
| Mora o.fl. 2011 1 | Chapman 2009 2 | Groombridge & Jenkins 2002 3 | ||||
|---|---|---|---|---|---|---|
| Lýstar | Spáð | Lýstar | Spáð | Lýstar | Spáð | |
| Dýr | 1.124.516 | 9.920.000 | 1.424.153 | 6.836.330 | 1.225.500 | 10.820.000 |
| Krómistar | 17.892 | 34.900 | 25.044 | 200.500 | — | — |
| Sveppir | 44.368 | 616.320 | 98.998 | 1.500.000 | 72.000 | 1.500.000 |
| Plöntur | 224.244 | 314.600 | 310.129 | 390.800 | 270.000 | 320.000 |
| Frumdýr | 16.236 | 72.800 | 28.871 | 1.000.000 | 80.000 | 600.000 |
| Dreifkjörnungar | — | — | 10.307 | 1.000.000 | 10.175 | — |
| Samtals | 1.438.769 | 10.960.000 | 1.897.502 | 10.897.630 | 1.657.675 | 13.240.000 |
Til eru ýmis verkefni sem miða að því að skrá lýstar tegundir á aðgengilegan hátt og internetið auðveldar þá vinnu. Engu að síður hefur verið bent á að miðað við núverandi hraða nýrra tegundalýsinga, 17.000 til 20.000 nýjar tegundir á ári samkvæmt State of Observed Species Report, muni taka nærri 500 ár að ljúka við að lýsa lífi á þessari jörð. Með tímanum verður verkefnið bæði erfiðara og auðveldara eftir því sem útdauði fjarlægir tegundir af jörðinni.
Að nefna og telja tegundir kann að virðast lítilvæg iðja í ljósi annarra þarfa mannkyns, en að ákvarða líffræðilega fjölbreytni er ekki einfaldlega bókhald yfir tegundir. Að lýsa tegund er flókið ferli þar sem líffræðingar ákvarða einstök einkenni lífveru og hvort sú lífvera tilheyri einhverri annarri lýstri tegund eða ættkvísl. Það gerir líffræðingum kleift að finna og þekkja tegundina eftir upphaflega uppgötvun og gerir þeim kleift að fylgja eftir spurningum um líffræði hennar. Að auki gera einstök einkenni hverrar tegundar hana mögulega verðmæta fyrir menn eða aðrar tegundir sem menn eru háðir.
Mynstur líffræðilegrar fjölbreytni
Líffræðilegri fjölbreytni er ekki jafnt dreift um jörðina. Í Viktoríuvatni voru næstum 500 tegundir síkliða einar og sér, ef litið er fram hjá öðrum fiskaættum í vatninu. Allar þessar tegundir fundust aðeins í Viktoríuvatni; þess vegna voru þessar 500 síkliðutegundir einlendar. Einlendar tegundir finnast aðeins á einum stað. Einlendar tegundir með mjög takmarkaða útbreiðslu eru sérstaklega viðkvæmar fyrir útdauða. Hærri flokkunarfræðileg stig, svo sem ættkvíslir og ættir, geta einnig verið einlend. Michigan-vatn hefur um 79 fisktegundir og margar þeirra finnast einnig í öðrum vötnum í Norður-Ameríku. Hvað skýrir muninn á fiskafjölbreytni þessara tveggja vatna? Viktoríuvatn er fornt hitabeltisvatn, en Michigan-vatn er nýlegt stöðuvatn á tempraða beltinu. Michigan-vatn í núverandi mynd er aðeins um 7.000 ára gamalt, en Viktoríuvatn í núverandi mynd er um 15.000 ára gamalt, þótt dæld þess sé um 400.000 ára gömul. Líflandafræðingar hafa bent á að þessir tveir þættir, breiddargráða og aldur, séu tvær af nokkrum tilgátum sem skýra mynstur líffræðilegrar fjölbreytni á jörðinni.
Starfstengsl
Líflandafræðingur
Líflandafræði er rannsókn á útbreiðslu tegunda heimsins – bæði í fortíð og nútíð. Starf líflandafræðinga er mikilvægt til að skilja efnislegt umhverfi okkar, hvernig umhverfið hefur áhrif á tegundir og hvernig umhverfisbreytingar hafa áhrif á útbreiðslu tegundar; það hefur einnig verið mikilvægt fyrir þróun nútíma þróunarkenninga. Líflandafræðingar þurfa að hafa skilning á bæði líffræði og vistfræði. Þeir þurfa einnig að vera vel að sér í þróunarfræðum, jarðvegsfræði og loftslagsfræði.
Það eru þrjú meginsvið rannsókna undir yfirskriftinni líflandafræði: vistfræðileg líflandafræði, söguleg líflandafræði (kölluð fornlíflandafræði) og verndunarlíflandafræði. Vistfræðileg líflandafræði rannsakar þá núverandi þætti sem hafa áhrif á útbreiðslu plantna og dýra. Söguleg líflandafræði, eins og nafnið gefur til kynna, rannsakar útbreiðslu tegunda í fortíðinni. Verndunarlíflandafræði einbeitir sér hins vegar að verndun og endurheimt tegunda byggt á þekktum sögulegum og núverandi vistfræðilegum upplýsingum. Hvert þessara sviða tekur til bæði dýralandafræði og plöntulandafræði – útbreiðslu dýra og plantna í fortíð og nútíð.
Eitt elsta mynstur sem sést hefur í vistfræði er að tegundafjölbreytni í næstum öllum flokkunarfræðilegum hópum eykst eftir því sem breiddargráða lækkar. Með öðrum orðum eykst líffræðileg fjölbreytni nær miðbaug (mynd 47.3).
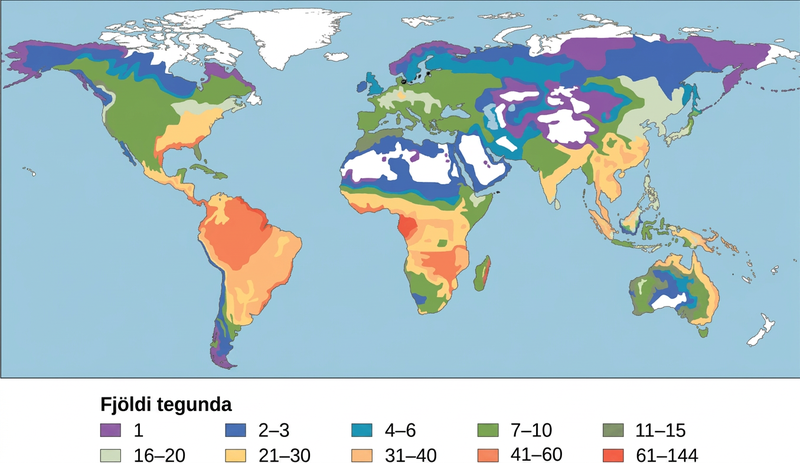
Ekki er enn ljóst hvers vegna líffræðileg fjölbreytni eykst nær miðbaug, en vísindamenn hafa sett fram nokkrar tilgátur. Einn þáttur gæti verið meiri aldur vistkerfa í hitabeltinu en á tempruðum svæðum; tempruðu svæðin voru að mestu lífvana eða líf þar dróst verulega saman á síðasta jökulskeiði. Hugmyndin er sú að meiri aldur gefi lengri tíma fyrir tegundamyndun. Önnur hugsanleg skýring er meiri bein sólarorka í hitabeltinu en á tempruðum svæðum og heimskautasvæðum. Flækjustig hitabeltisvistkerfa gæti ýtt undir tegundamyndun með því að auka misleitni, það er fjölda vistsessa, miðað við hærri breiddargráður. Meiri misleitni veitir fleiri tækifæri til samþróunar, sérhæfingar og ef til vill meiri valþrýstings sem leiðir til aðgreiningar stofna. Þessi tilgáta glímir þó við ákveðna hringröksemd: vistkerfi með fleiri tegundum ýta undir tegundamyndun, en hvernig fengu þau fleiri tegundir til að byrja með?
Litið hefur verið svo á að hitabeltið sé stöðugra en tempruð svæði, sem hafa áberandi árstíðabundinn breytileika í loftslagi og daglengd. Hitabeltið hefur sínar eigin tegundir af árstíðabundnum sveiflum, svo sem úrkomu, en almennt er gert ráð fyrir að þar séu stöðugri umhverfisaðstæður og þessi stöðugleiki gæti ýtt undir tegundamyndun í mjög sérhæfða sessa.
Hvaða ferlar sem liggja að baki er ljóst að öll stig líffræðilegrar fjölbreytni eru mest í hitabeltinu. Þar er hlutfall einlendra tegunda einnig hæst og þar eru fleiri heitir reitir líffræðilegrar fjölbreytni. Þessi mikla fjölbreytni þýðir þó líka að þekking á tegundum er því miður mjög lítil og hættan á tapi líffræðilegrar fjölbreytni er mikil.
Verndun líffræðilegrar fjölbreytni
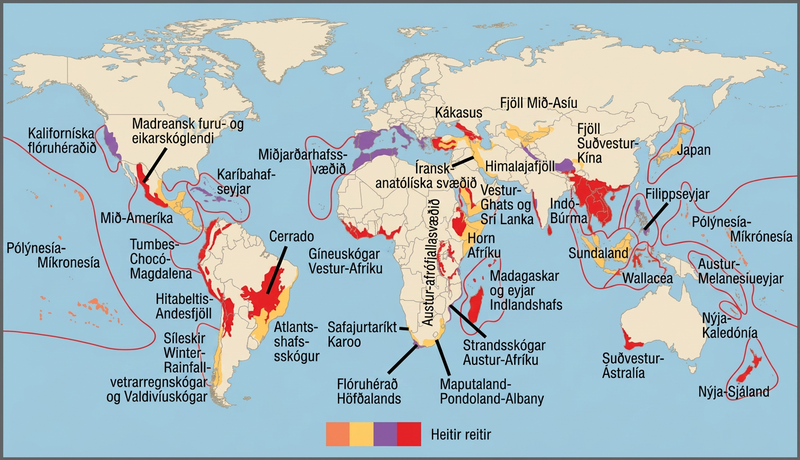
Árið 1988 þróaði breski umhverfisverndarsinninn Norman Myers verndunarhugtak til að bera kennsl á svæði sem eru auðug af tegundum og í verulegri hættu á tegundatapi: heita reiti líffræðilegrar fjölbreytni. Heitir reitir líffræðilegrar fjölbreytni eru landfræðileg svæði með mikinn fjölda einlendra tegunda. Tilgangur hugtaksins var að finna mikilvæg svæði fyrir verndunaraðgerðir, eins konar forgangsröðun náttúruverndar. Með því að vernda heita reiti geta stjórnvöld verndað meiri fjölda tegunda. Upprunalegu viðmiðin fyrir heitan reit voru að þar væru 1.500 eða fleiri einlendar plöntutegundir og að 70 prósent svæðisins hefðu raskast af mannavöldum. Nú eru til 34 heitir reitir líffræðilegrar fjölbreytni (mynd 47.4) sem innihalda mikinn fjölda einlendra tegunda, þar á meðal helming allra einlendra plantna jarðar.
Breytingar á líffræðilegri fjölbreytni í gegnum jarðsöguna
Fjöldi tegunda á jörðinni, eða á hvaða landfræðilega svæði sem er, er afleiðing af jafnvægi tveggja þróunarferla sem eru stöðugt í gangi: tegundamyndunar og útdauða. Hvort tveggja eru náttúruleg „fæðingar-“ og „dauða-“ ferli stórþróunar. Þegar hraði tegundamyndunar fer að verða meiri en hraði útdauða mun tegundum fjölga; sömuleiðis mun tegundum fækka þegar hraði útdauða fer að verða meiri en hraði tegundamyndunar. Í gegnum sögu jarðar hafa þessir tveir ferlar sveiflast – sem hefur stundum leitt til stórkostlegra breytinga á fjölda tegunda á jörðinni eins og sést í steingervingasögunni (mynd 47.5).
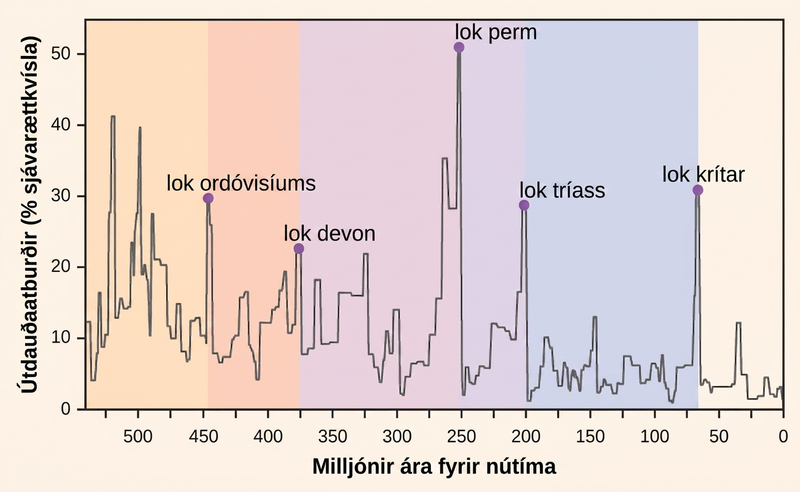
Steingervingafræðingar hafa greint fimm jarðlög í steingervingasögunni sem virðast sýna skyndilegt og stórkostlegt tap á líffræðilegri fjölbreytni: 25 til 55 prósent ættkvísla og meira en helmingur allra þáverandi tegunda hverfur úr steingervingasögunni. Slíkir atburðir kallast fjöldaútdauðar. Til eru margir minni, en samt stórkostlegir, útdauðaatburðir, en fjöldaútdauðarnir fimm hafa fengið mesta athygli rannsakenda. Færa má rök fyrir því að þeir séu aðeins fimm öfgafyllstu atburðirnir í samfelldri röð stórra útdauðaatburða á öld lífsins, það er síðustu 542 milljón árin. Í flestum tilfellum eru tilgátur um orsakir enn umdeildar; nýjasti fjöldaútdauðinn virðist þó skýrari.
Steingervingasaga fjöldaútdauðanna var grundvöllurinn fyrir skilgreiningu tímabila í jarðsögunni, svo þeir eiga sér yfirleitt stað við skil milli jarðsögutímabila. Breytingin á steingervingum frá einu tímabili til annars endurspeglar stórkostlegt tap tegunda og hægfara uppruna nýrra tegunda. Þessi skil má sjá í berglögum. Tafla 47.2 veitir gögn um fjöldaútdauðana fimm.
Fjöldaútdauðarnir fimm
| Jarðsögutímabil | Heiti fjöldaútdauða | Tími (fyrir milljónum ára) |
|---|---|---|
| Ordóvisíum–Sílúr | lok ordóvisíum O–S | 450–440 |
| Síð-Devon | lok Devon | 375–360 |
| Perm–Trías | lok Perm | 251 |
| Trías–Júra | lok Trías | 205 |
| Krít–Paleógen | lok Krítar K-Pg (K–T) | 65,5 |
Útdauðinn á mörkum ordóvisíum og sílúr er fyrsti skráði fjöldaútdauðinn og sá næststærsti. Á þessu tímabili dóu um 85 prósent sjávartegunda út, en fáar tegundir lifðu utan hafsins. Helsta tilgátan um orsök hans er jökulskeið og síðan hlýnun. Útdauðinn samanstendur í raun af tveimur útdauðaatburðum sem aðskildir voru með um einni milljón ára. Fyrri atburðurinn stafaði af kólnun og sá síðari af hlýnuninni sem fylgdi í kjölfarið. Loftslagsbreytingarnar höfðu áhrif á hitastig og sjávarstöðu. Sumir vísindamenn hafa bent á að gammablossi frá nálægri sprengistjörnu gæti hafa valdið útdauðanum á mörkum ordóvisíum og sílúr. Gammablossinn hefði getað eytt verndandi ósonlagi jarðar og hleypt mikilli útfjólublárri geislun frá sólinni niður á yfirborðið, sem gæti skýrt loftslagsbreytingarnar á þessum tíma. Tilgátan er mjög ágiskunarkennd og áhrif utan úr geimnum á sögu jarðar eru virkt rannsóknarefni. Endurheimt líffræðilegrar fjölbreytni eftir fjöldaútdauðann tók 5 til 20 milljónir ára, eftir staðsetningu.
Útdauðinn á síð-Devon gæti hafa átt sér stað yfir tiltölulega langan tíma. Hann virðist hafa haft mest áhrif á sjávartegundir en ekki svo mjög á plöntur eða dýr sem lifðu á landi. Á þessum tíma dóu 75 prósent allra tegunda út. Orsakir þessa útdauða eru illa skilgreindar.
Útdauðinn við lok perm var sá mesti í sögu lífsins. Reyndar mætti færa rök fyrir því að jörðin hafi orðið næstum lífvana í þessum útdauðaatburði. Áætlað er að 96 prósent allra sjávartegunda og 70 prósent allra landtegunda hafi tapast. Það var á þessum tíma, til dæmis, sem þríbrotar, hópur sem lifði af útdauðann á mörkum ordóvisíum og sílúr, dóu út. Orsakir þessa fjöldaútdauða eru ekki ljósar, en helsta skýringin er talin vera langvarandi og útbreidd eldvirkni sem leiddi til stjórnlausrar hlýnunar jarðar. Hafið varð að mestu súrefnissnautt og það kæfði líf í sjónum. Það tók fjölbreytni landferfætlna 30 milljónir ára að jafna sig eftir útdauðann við lok perm. Útdauðinn við lok perm breytti verulega samsetningu líffræðilegrar fjölbreytni jarðar og framvindu þróunar.
Orsakir útdauðans á mörkum trías og júra eru ekki ljósar og vísindamenn deila um tilgátur, þar á meðal loftslagsbreytingar, árekstur smástirnis og eldgos. Útdauðaatburðurinn varð rétt áður en risameginlandið Pangea brotnaði upp, þótt nýlegar rannsóknir bendi til þess að útdauðinn hafi hugsanlega orðið hægar í gegnum trías. Á þessum tíma dóu 76 til 80 prósent allra tegunda út.
Orsakir útdauðans við lok krítar eru þær sem eru best skildar. Það var í þessum útdauðatburði fyrir um 65 milljónum ára sem meirihluti risaeðlanna, ríkjandi hryggdýrahópur í milljónir ára, hvarf af jörðinni (að undanskildum hópi þerapoda sem varð uppruni fugla). Á heildina litið dóu um 75 prósent allra tegunda út á þessu tímabili.
Nú er talið að orsök þessa útdauða hafi verið hrikalegur árekstur stórs loftsteins, eða smástirnis, undan ströndum þess sem nú er Júkatanskagi. Þessi tilgáta, sem fyrst var sett fram árið 1980, var róttæk skýring byggð á snörpum toppi í magni iridíums, sem berst inn í lofthjúp okkar frá loftsteinum á nokkuð jöfnum hraða en finnst annars ekki á yfirborði jarðar, í berglaginu sem markar mörkin milli krítar og paleógens (mynd 47.6). Þessi mörk sýna hvarf risaeðla úr steingervingasögunni, ásamt mörgum öðrum flokkunareiningum. Vísindamennirnir sem fundu iridíumtoppinn túlkuðu hann sem hraðt innflæði iridíums úr geimnum í lofthjúpinn, í formi stórs smástirnis, frekar en sem merki um hægari setmyndun á þessu tímabili. Þetta var róttæk skýring, en fundur árekstrargígs af réttum aldri og stærð árið 1991 gerði tilgátuna trúverðugri. Nú styður gnægð jarðfræðilegra gagna kenninguna. Endurheimtartími líffræðilegrar fjölbreytni eftir útdauðann við lok krítar er styttri, á jarðsögulegum tímakvarða, en eftir útdauðann við lok perm, eða um 10 milljónir ára.
Annar möguleiki, sem kann að hafa fallið saman við árekstur Júkatan-smástirnisins, var umfangsmikil eldvirkni sem hófst fyrir um 66 milljónum ára, um svipað leyti og áreksturinn, við lok krítar. Hraunflæðið þakti yfir 50 prósent af því sem nú er Indland. Losun eldfjallagasa, einkum brennisteinsdíoxíðs, í þessum umfangsmiklu gosum stuðlaði að loftslagsbreytingum sem gætu hafa komið fjöldaútdauðanum af stað.
Sjónræn tenging

Vísindamenn mældu hlutfallslegt magn burknagróa ofan og neðan við K-Pg mörkin í þessu bergsýni. Hver eftirfarandi fullyrðinga lýsir niðurstöðum þeirra líklegast?
Tengill í námsefni
Skoðaðu þessa gagnvirku vefsíðu um fjöldaútdauða.
Útdauðinn á pleistósen
Útdauðinn á pleistósen er einn af minni útdauðaatburðunum og jafnframt nýlegur. Vel er þekkt að stór hluti risadýralífs Norður-Ameríku, og að nokkru leyti Evrasíu, hvarf undir lok síðasta jökulskeiðs; þar var um að ræða stór hryggdýr. Útdauðinn virðist hafa orðið á tiltölulega afmörkuðu tímabili fyrir 10.000 til 12.000 árum. Í Norður-Ameríku var tapið mjög áberandi og náði meðal annars til loðfíla, þótt einangraður stofn hafi lifað á Wrangel-eyju í Kanada þar til fyrir um 4.000 árum, mastódonta, risabifra, risajarðletidýra, sverðtannakatta og norður-ameríska úlfaldans, svo fátt eitt sé nefnt. Í byrjun 20. aldar bentu vísindamenn fyrst á þann möguleika að ofveiði hefði valdið hröðum útdauða þessara stóru dýra. Rannsóknir á þessari tilgátu halda áfram.
Almennt séð fór tímasetning útdauðans á pleistósen saman við komu frummanna, ef til vill fyrir allt að 40.000 árum, en ekki við atburði tengda loftslagsbreytingum, sem er helsta samkeppnistilgátan fyrir þessum útdauða. Útdauðinn hófst í Ástralíu fyrir um 40.000 til 50.000 árum, rétt eftir komu manna á svæðið: pokaljón, risavaxinn eins tonns vombati og nokkrar tegundir risakengúra hurfu. Í Norður-Ameríku varð útdauði næstum allra stórra spendýra fyrir 10.000–12.000 árum. Þau einu sem eftir eru eru minni spendýr eins og birnir, vapítihirtir, elgir og fjallaljón. Að lokum, á mörgum afskekktum úthafseyjum, átti útdauði margra tegunda sér stað samhliða komu manna. Ekki voru stór dýr á öllum eyjunum, en þar sem stór dýr voru til staðar voru þau oft þvinguð til útdauða. Madagaskar var numið landi fyrir um 2,000 árum og stóru spendýrin sem þar lifðu dóu út. Evrasía og Afríka sýna ekki þetta mynstur, en þar komu safnarar og veiðimenn heldur ekki nýlega til sögunnar. Þvert á móti komu menn til Evrasíu fyrir hundruðum þúsunda ára. Þetta efni er enn vettvangur virkra rannsókna og tilgátusmíða. Það virðist ljóst að jafnvel þótt loftslag hafi spilað inn í, þá hafi veiðar manna í flestum tilfellum hrint útdauðanum af stað.
Sjötti fjöldaútdauðinn, eða fjöldaútdauði nútíma á hólósen, virðist hafa hafist fyrr en áður var talið og stafar að mestu af raskandi athöfnum nútímamanna, Homo sapiens. Frá upphafi hólósen eru fjölmörg dæmi um nýlegan útdauða einstakra tegunda sem varðveitt eru í ritheimildum manna. Flest þeirra falla saman við útþenslu evrópskra nýlenda frá 16. öld.
Eitt af eldri og þekktari dæmunum er dódófuglinn. Þessi einkennilegi dúfulíki fugl lifði í skógum Máritíus, eyju í Indlandshafi, og dó út um 1662. Dódófuglinn var veiddur til matar af sjómönnum og var auðveld bráð því hann nálgaðist fólk óttalaus; hann hafði ekki þróast í návist manna. Svín, rottur og hundar sem evrópsk skip fluttu til eyjarinnar drápu einnig unga og egg dódófuglsins.
Stellers-sækýrin dó út árið 1768. Hún var skyld manatíum og lifði líklega eitt sinn meðfram norðvesturströnd Norður-Ameríku. Evrópubúar fundu Stellers-sækúna fyrst árið 1741 og hún var ofveidd vegna kjöts og lýsis. Síðasta dýrið var drepið árið 1768. Aðeins 27 ár liðu frá fyrstu kynnum Evrópubúa af tegundinni þar til hún dó út.
Síðan 1900 hafa ýmsar tegundir dáið út, þar á meðal eftirfarandi:
Þetta eru aðeins fáeinir af þeim útdauðum sem skráðir hafa verið á síðustu 500 árum. Alþjóðanáttúruverndarsamtökin (IUCN) halda skrá yfir útdauðar tegundir og tegundir í útrýmingarhættu sem kallast Rauði listinn. Listinn er ekki tæmandi, en hann lýsir 380 útdauðum hryggdýrategundum eftir árið 1500, þar af voru 86 reknar til útdauða með ofveiði eða ofnýtingu.
Áætlanir um útdauðahraða nútímans
Áætlanir um útdauðatíðni eru torveldaðar af þeirri staðreynd að flestir útdauðar eiga sér líklega stað án þess að eftir því sé tekið. Líklegt er að menn taki eftir útdauða fugls eða spendýrs, sérstaklega ef dýrið hefur verið veitt eða nýtt á annan hátt. En það eru margar lífverur sem vekja minni áhuga manna (ekki endilega minna virði) og margar sem eru ólýstar.
Áætlað er að bakgrunnsútdauðatíðni sé um það bil ein tegund á hverja milljón tegunda á ári (E/MSY). Til dæmis, ef gert er ráð fyrir að um tíu milljónir tegunda séu til, eru væntingarnar þær að tíu tegundir deyi út á hverju ári.
Ein nútímaáætlun um útdauðatíðni notar útdauða í ritheimildum frá árinu 1500. Fyrir fugla eina og sér gefur þessi aðferð áætlun upp á 26 E/MSY. Hins vegar gæti þetta gildi verið vanmat af þremur ástæðum. Í fyrsta lagi hefðu margar tegundir ekki verið lýstar fyrr en mun síðar á tímabilinu, svo tap þeirra hefði farið framhjá mönnum. Í öðru lagi fer fjöldi nýlega útdauðra hryggdýrategunda vaxandi vegna þess að útdauðum tegundum er nú lýst út frá beinaleifum. Og í þriðja lagi eru sumar tegundir líklega þegar útdauðar þótt náttúruverndarsinnar séu tregir til að nefna þær sem slíkar. Þegar tekið er tillit til þessara þátta hækkar áætluð útdauðatíðni nær 100 E/MSY. Spáð er að tíðnin í lok aldarinnar verði 1.500 E/MSY.
Önnur nálgun við að áætla núverandi útdauðatíðni er að tengja tap tegunda við tap búsvæða með því að mæla tap skógarsvæða og skilja sambandið milli tegundafjölda og svæðis. Samband tegundafjölda og svæðis vísar til þess hraða sem nýjar tegundir koma fram þegar stærð rannsakaðs svæðis er aukin. Rannsóknir hafa sýnt að fjöldi tegunda sem eru til staðar eykst eftir því sem stærð eyjar eykst. Sýnt hefur verið fram á að þetta fyrirbæri eigi einnig við um önnur eyjulík búsvæði, svo sem tepui-fjöllin í Venesúela, sem eru umkringd regnskógi. Ef þessu sambandi er snúið við, þá mun tegundum sem þar lifa fækka ef búsvæðið minnkar. Áætlanir um útdauðatíðni byggðar á tapi búsvæða og sambandi tegundafjölda og svæðis hafa bent til þess að með um 90 prósenta tapi búsvæða megi búast við að 50 prósent tegunda deyi út. Áætlanir byggðar á tegunda-svæðis sambandi hafa leitt til útreikninga á útdauðatíðni tegunda sem nemur um 1000 E/MSY og hærra. Almennt séð sýna raunverulegar athuganir ekki þetta magn taps og bent hefur verið á að seinkun verði á útdauða. Nýleg vinna hefur einnig dregið í efa notagildi tegunda-svæðis sambandsins við mat á tapi tegunda. Þessi vinna færir rök fyrir því að tegunda-svæðis sambandið leiði til ofmats á útdauðatíðni. Betra samband til að nota gæti verið samband einlendra tegunda og svæðis (endemics-area relationship). Notkun þessarar aðferðar myndi lækka áætlanir niður í um 500 E/MSY á komandi öld. Athugið að þetta gildi er enn 500 sinnum hærra en bakgrunnsútdauðatíðnin.
