23.1 Uppruni heilkjörnunga
23.1 Uppruni heilkjörnunga
Námsmarkmið
Í lok þessa hluta muntu geta gert eftirfarandi:
Lífverur eru flokkaðar í þrjú lén: Fornbakteríur, Bakteríur og Heilkjörnungar. Fyrstu tvær þróunarlínurnar ná yfir allar dreifkjörnungafrumur og sú þriðja inniheldur alla heilkjörnunga. Mjög fátæklegar steingervingaheimildir koma í veg fyrir að við getum ákvarðað hvernig fyrstu meðlimir hverrar þessara þróunarlína litu út, svo það er mögulegt að allir atburðirnir sem leiddu til síðasta sameiginlega forföður núlifandi heilkjörnunga verði áfram óþekktir. Hins vegar veitir samanburðarlíffræði núlifandi lífvera og hinar takmörkuðu steingervingaheimildir nokkra innsýn í þróun heilkjörnunga.
Elstu steingervingar sem fundist hafa virðast vera af léni baktería. Þeir eru um 3,5 til 3,8 milljarða ára gamlir og eru þekkjanlegir vegna tiltölulega flókinnar byggingar og, miðað við dreifkjörnunga, tiltölulega stórra frumna. Flestir aðrir dreifkjörnungar hafa litlar frumur, 1 eða 2 µm að stærð, og erfitt væri að greina þær sem steingervinga. Steingerðir stromatólítar benda til þess að að minnsta kosti sumir dreifkjörnungar hafi lifað í samfélögum með víxlverkun, og vísbendingar frá byggingu núlifandi heilkjörnungafrumna benda til þess að svipað samspil forfeðra hafi átt þátt í uppruna heilkjörnunga. Flestir núlifandi heilkjörnungar hafa frumur sem mælast 10 µm eða stærri. Byggingar af þessari stærð, sem gætu verið steingerðar leifar snemmbærra heilkjörnunga, koma fram í jarðsögunni í setlögum sem eru um 2,1 milljarðs ára gömul.
Einkenni heilkjörnunga
Gögn frá þessum steingervingum, sem og frá rannsóknum á erfðamengjum núlifandi lífvera, hafa leitt samanburðarlíffræðinga til þeirrar niðurstöðu að núlifandi heilkjörnungar séu allir afkomendur eins sameiginlegs forföður. Kortlagning á einkennum sem finnast í öllum helstu hópum heilkjörnunga leiðir í ljós að eftirfarandi einkenni eru til staðar í að minnsta kosti sumum meðlimum hverrar helstu þróunarlínu, eða á einhverjum hluta lífsferils þeirra, og hljóta því að hafa verið til staðar hjá síðasta sameiginlega forföðurnum.
Innra samlífi og þróun heilkjörnunga
Áður en við ræðum uppruna heilkjörnunga er fyrst mikilvægt að skilja að allir núlifandi heilkjörnungar eru líklega afkomendur blendingslífveru sem var samsett úr hýsilfrumu og einni eða fleiri frumum alfa-próteóbaktería sem tóku sér bólfestu inni í henni. Þetta meginþema í uppruna heilkjörnunga er þekkt sem innra samlífi, þar sem ein fruma gleypir aðra þannig að gleypta fruman lifir af og báðar frumurnar hagnast. Yfir margar kynslóðir getur samlífssamband leitt til tveggja lífvera sem eru svo algerlega háðar hvor annarri að hvorug gæti lifað af ein. Atburðir tengdir innra samlífi stuðluðu líklega að uppruna síðasta sameiginlega forföður heilkjörnunga nútímans og að síðari fjölbreytni í ákveðnum þróunarlínum heilkjörnunga (Mynd 23.5). Svipuð tengsl byggð á innra samlífi eru ekki óalgeng hjá núlifandi heilkjörnungum. Áður en þetta er útskýrt nánar er nauðsynlegt að huga að efnaskiptum hjá dreifkjörnungum.
Efnaskipti dreifkjörnunga
Mörg mikilvæg efnaskiptaferli komu fram hjá dreifkjörnungum; hins vegar finnast sum þessara ferla, eins og niturbinding, aldrei hjá heilkjörnungum. Ferli loftháðrar öndunar finnst í öllum helstu þróunarlínum heilkjörnunga og það fer fram í hvatberunum. Loftháð öndun finnst einnig í mörgum þróunarlínum dreifkjörnunga, en hún er ekki til staðar í þeim öllum, og miklar vísbendingar benda til þess að slíkir loftfirrtir dreifkjörnungar hafi aldrei stundað loftháða öndun og ekki heldur forfeður þeirra.
Þó að andrúmsloft nútímans sé um 20 prósent sameindasúrefni (O2), sýna jarðfræðilegar vísbendingar að upphaflega hafi O2 vantað. Án súrefnis væri ekki búist við loftháðri öndun og lifandi verur hefðu reitt sig á loftfirrta öndun eða gerjun í staðinn. Á einhverjum tímapunkti fyrir 3,2 til 3,5 milljörðum ára byrjuðu sumir dreifkjörnungar að nota orku frá sólarljósi til að knýja uppbyggingarferli sem afoxa koldíoxíð til að mynda lífræn efnasambönd. Það er að segja, þeir þróuðu með sér hæfileikann til ljóstillífunar. Vetni, fengið úr ýmsum uppsprettum, var „fangað“ með ljósknuðum efnahvörfum til að afoxa bundið koldíoxíð í Calvin-hringnum. Hópur Gram-neikvæðra baktería sem gaf af sér blágerla notaði vatn sem vetnisgjafa og losaði O2 sem úrgangsefni fyrir um 2,2 milljörðum ára.
Að lokum safnaðist magn súrefnis frá ljóstillífun upp í sumum umhverfum í það mikinn mæli að það ógnaði lifandi lífverum, þar sem það getur skemmt mörg lífræn efnasambönd. Ýmis efnaskiptaferli þróuðust sem vernduðu lífverur gegn súrefni, og eitt þeirra, loftháð öndun, myndaði einnig mikið magn af ATP. Það varð útbreitt meðal dreifkjörnunga, þar á meðal í frílifandi hópi sem við köllum nú alfa-próteóbakteríur. Lífverur sem tileinkuðu sér ekki loftháða öndun urðu að halda sig í súrefnislausu umhverfi. Upphaflega voru súrefnisrík umhverfi líklega staðbundin í kringum staði þar sem blágerlar voru algengir og virkir, en fyrir um 2 milljörðum ára sýna jarðfræðilegar vísbendingar að súrefni var farið að safnast upp í meiri styrk í andrúmsloftinu. Súrefnisstyrkur svipaður því sem er í dag kom aðeins fram á síðustu 700 milljónum ára.
Rifjaðu upp að fyrstu steingervingarnir sem við teljum vera heilkjörnunga eru um 2 milljarða ára gamlir, svo þeir virðast hafa þróast og orðið fjölbreyttir hratt þegar súrefnisstig jókst. Rifjaðu einnig upp að allir núlifandi heilkjörnungar eru komnir af forföður sem hafði hvatbera. Þessi frumulíffæri sáust fyrst í ljóssmásjá seint á 19. öld, þar sem þau virtust vera nokkuð ormlaga byggingar sem virtust hreyfa sig um í frumunni. Sumir snemmbúnir athugendur lögðu til að þetta gætu verið bakteríur sem lifðu inni í hýsilfrumum, en þessar tilgátur voru óþekktar eða þeim hafnað í flestum vísindasamfélögum.
Kenningin um innra samlífi
Eftir því sem frumulíffræði þróaðist á tuttugustu öldinni varð ljóst að hvatberar voru frumulíffærin sem bera ábyrgð á framleiðslu ATP með loftháðri öndun, þar sem súrefni var lokarafeindaþegi. Á sjöunda áratugnum þróaði bandaríski líffræðingurinn Lynn Margulis við Boston-háskóla innri samlífskenninguna, sem segir að heilkjörnungar gætu hafa orðið til við það að ein fruma innlimaði aðra, önnur lifði inni í hinni, og þær samþróuðust með tímanum þar til frumurnar voru ekki lengur þekkjanlegar sem aðskildar frumur og deildu erfðafræðilegri stjórn á gagnkvæmu efnaskiptaferli til að framleiða ATP. Árið 1967 kynnti Margulis ný gögn til stuðnings vinnu sinni við kenninguna og rökstuddi niðurstöður sínar með örverufræðilegum sönnunargögnum. Þótt verkum Margulis hafi í fyrstu verið mætt með andstöðu, er þessi grunnþáttur þessarar einu sinni byltingarkenndu tilgátu nú almennt viðurkenndur, og vinna heldur áfram við að afhjúpa skrefin í þessu þróunarferli og lykilþátttakendurna.
Þó að efnaskiptafrumulíffærin og genin sem bera ábyrgð á mörgum orkuöflunarferlum virðist eiga uppruna sinn í bakteríum, virðast kjarnagen okkar og sameindavélarnar sem bera ábyrgð á eftirmyndun og tjáningu vera skyldari þeim sem finnast í fornbakteríum. Margt á enn eftir að skýra varðandi hvernig þetta samband kom til; þetta heldur áfram að vera spennandi uppgötvunarsvið í líffræði. Til dæmis er ekki vitað hvort atburðurinn sem leiddi til innra samlífis hvatbera átti sér stað áður eða eftir að hýsilfruman fékk kjarna. Slíkar lífverur væru meðal útdauðra forvera síðasta sameiginlega forföður heilkjörnunga.
Hvatberar
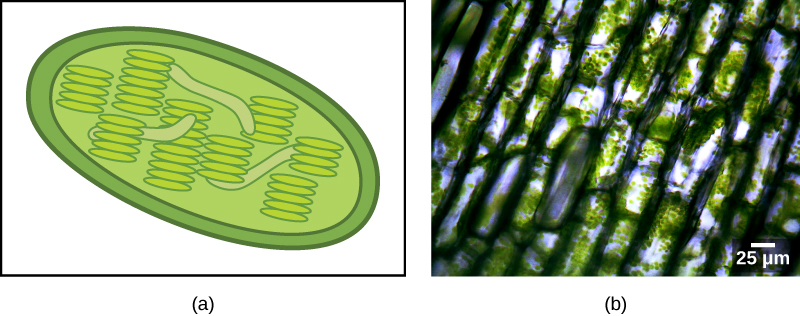
Eitt af helstu einkennunum sem greinir dreifkjörnunga frá heilkjörnungum er tilvist hvatbera, eða smækkaðra afleiðna þeirra, í nánast öllum heilkjörnungafrumum. Heilkjörnungafrumur geta innihaldið allt frá einum upp í nokkur þúsund hvatbera, allt eftir orkunotkun frumunnar; í mönnum eru þeir algengastir í lifur og beinagrindarvöðvum. Hver hvatberi mælist 1 til 10 eða fleiri míkrómetrar að lengd og er til staðar í frumunni sem frumulíffæri sem getur verið egglaga, ormlaga eða flókið greint (Mynd 23.2). Hins vegar, þó að þeir kunni að hafa átt uppruna sinn sem frjálslifandi loftháðar lífverur, geta hvatberar ekki lengur lifað af og fjölgað sér utan frumunnar.
Hvatberar hafa nokkra eiginleika sem benda til skyldleika þeirra við alfa-próteóbakteríur (Mynd 23.5). Alfa-próteóbakteríur eru stór hópur baktería sem inniheldur tegundir í samlífi við plöntur, sýkla sem geta borist í menn með mítlum, og margar frjálslifandi tegundir sem nota ljós til orku. Hvatberar hafa sín eigin erfðamengi, með hringlaga litningi sem er festur við innri himnuna. Hvatberar hafa einnig sérstök ríbósóm og flutnings-RNA sem líkjast sömu þáttum í dreifkjörnungum. Athyglisverður eiginleiki hvatbera er að margir þeirra sýna minniháttar frávik frá hinum almenna erfðakóða. Hins vegar eru mörg genanna fyrir öndunarprótein nú staðsett í kjarnanum. Þegar þessi gen eru borin saman við gen annarra lífvera virðast þau vera af uppruna alfa-próteóbaktería. Í sumum hópum heilkjörnunga finnast slík gen í hvatberunum, en í öðrum hópum finnast þau í kjarnanum. Þetta hefur verið túlkað sem sönnun þess að á þróunartíma hafi gen flust frá litningi innri samlífislífverunnar yfir í erfðamengi hýsilsins. Þetta augljósa „tap“ á genum hjá innri samlífislífverunni er líklega ein skýring þess að hvatberar geta ekki lifað án hýsils.
Önnur sönnunargögn sem styðja þá hugmynd að hvatberar séu komnir af innra samlífi koma frá byggingu hvatberans sjálfs. Flestir hvatberar eru í laginu eins og alfa-próteóbakteríur og eru umkringdir tveimur himnum; innri himnan er bakteríukyns í eðli sínu en ytri himnan er heilkjörnungakyns. Þetta er nákvæmlega það sem búast mætti við ef ein himnubundin lífvera væri innlimuð í bólu af annarri himnubundinni lífveru. Ytri hvatberahimnan varð til úr bólunni sem umlukti innri samlífislífveruna, en innri himnan varð til úr frumuhimnu hennar. Innri himna hvatberans er umfangsmikil og myndar verulegar innfellingar sem líkjast áferðarmiklu yfirborði alfa-próteóbaktería. Mergurinn og innri himnan eru rík af ensímum sem eru nauðsynleg fyrir loftháða öndun.

Þriðja línan af sönnunargögnum kemur frá myndun nýrra hvatbera. Hvatberar skipta sér sjálfstætt með ferli sem líkist tvískiptingu hjá dreifkjörnungum. Hvatberar verða eingöngu til af fyrri hvatberum; þeir eru ekki myndaðir frá grunni af heilkjörnungafrumunni. Hvatberar geta runnið saman; og þeir geta færst til innan frumunnar vegna samspils við frumugrindina. Þeir fjölga sér innan frumunnar sem umlykur þá og dreifast með umfryminu þegar fruma skiptir sér eða tvær frumur renna saman. Þess vegna, þó að þessi frumulíffæri séu mjög samofin heilkjörnungafrumunni, fjölga þau sér enn eins og þau væru sjálfstæðar lífverur innan frumunnar. Hins vegar er fjölgun þeirra samstillt við virkni og skiptingu frumunnar. Þessir eiginleikar styðja allir þá kenningu að hvatberar hafi eitt sinn verið frjálslifandi dreifkjörnungar.
Sumir núlifandi heilkjörnungar eru loftfirrtir og geta ekki lifað af í of miklu súrefni. Hins vegar virðast fáeinir skorta frumulíffæri sem hægt væri að þekkja sem hvatbera. Á áttunda áratugnum og fram á snemma á tíunda áratugnum lögðu margir líffræðingar til að sumir þessara heilkjörnunga væru komnir af forfeðrum þar sem þróunarlínurnar hefðu aðskilist frá þróunarlínu hvatberaberandi heilkjörnunga áður en innra samlífi átti sér stað. Síðari niðurstöður benda til þess að smækkuð frumulíffæri finnist í flestum, ef ekki öllum, loftfirrtum heilkjörnungum, og að nánast allir heilkjörnungar virðist bera einhver gen í kjörnum sínum sem eru af hvatberauppruna.
Auk loftháðrar framleiðslu á ATP, hafa hvatberar nokkur önnur efnaskiptahlutverk. Eitt þessara hlutverka er að mynda klasa af járni og brennisteini sem eru mikilvægir hjálparþættir margra ensíma. Slík hlutverk eru oft tengd hinum smækkuðu frumulíffærum af hvatberauppruna í loftfirrtum heilkjörnungum. Frumveran Monocercomonoides, sem lifir í meltingarvegi hryggdýra, virðist vera undantekning; hún hefur enga hvatbera og erfðamengi hennar inniheldur hvorki gen sem eru komin frá hvatberum né kjarnagen sem tengjast viðhaldi hvatbera. Hún er hins vegar skyld öðrum frumverum með smækkaða hvatbera og táknar líklega endapunkt í smækkun hvatbera. Þó að flestir líffræðingar samþykki að síðasti sameiginlegi forfaðir heilkjörnunga hafi haft hvatbera, virðist sem hið flókna samband milli hvatbera og hýsilfrumu þeirra haldi áfram að þróast.
Plastíð
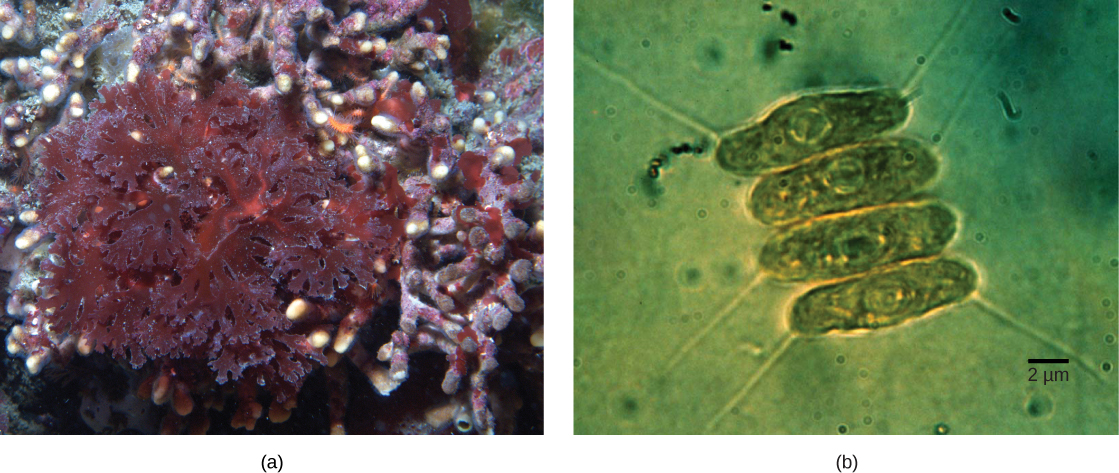
Sumir hópar heilkjörnunga eru ljóstillífandi. Frumur þeirra innihalda, auk hefðbundinna frumulíffæra heilkjörnunga, aðra tegund frumulíffæra sem kallast plastíð. Þegar slíkar frumur stunda ljóstillífun eru plastíð þeirra rík af litarefninu blaðgrænu a og ýmsum öðrum litarefnum, kölluðum hjálparlitarefni, sem taka þátt í að beisla orku úr ljósi. Ljóstillífandi plastíð kallast grænukorn (Mynd 23.3).
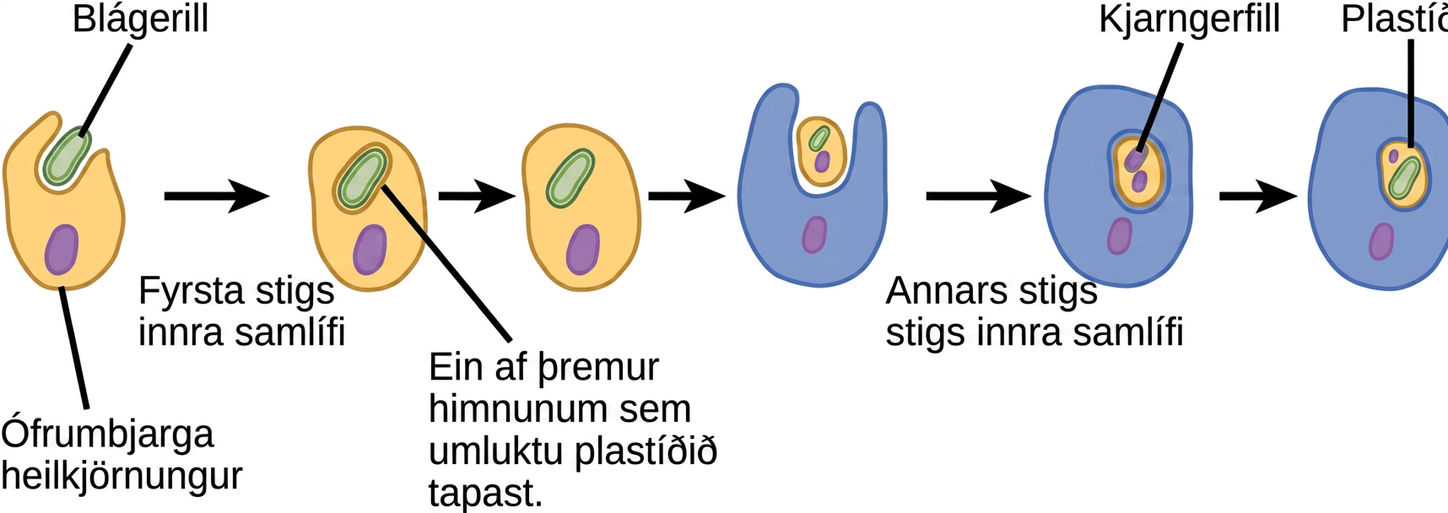
Líkt og hvatberar virðast plastíð eiga uppruna sinn í innra samlífi. Þessi tilgáta var einnig lögð fram og varin með fyrstu beinu sönnunargögnunum af Lynn Margulis. Við vitum nú að plastíð eru komin af blágerlum sem lifðu inni í frumum forföður sem var loftháður, ófrumbjarga heilkjörnungur. Þetta kallast fyrsta stigs innra samlífi og plastíð af frumuppruna eru umkringd tveimur himnum. Hins vegar eru bestu sönnunargögnin þau að upptaka blágerla sem innri samlífislífverur hafi gerst tvisvar í sögu heilkjörnunga. Í öðru tilvikinu tók sameiginlegur forfaðir stóra hópsins Archaeplastida upp blágeril sem innri samlífislífveru; í hinu tilvikinu tók forfaðir hinnar litlu amöbulaga Rhizaria-flokkunareiningar, Paulinella, upp annan blágeril sem innri samlífislífveru. Nánast allir ljóstillífandi heilkjörnungar eru komnir af fyrri atburðinum, og aðeins örfáar tegundir eru komnar af hinum, sem í þróunarlegu tilliti virðist vera nýlegri.
Blágerlar eru hópur Gram-neikvæðra baktería sem búa yfir öllum hefðbundnum byggingareiginleikum hópsins. Hins vegar, ólíkt flestum dreifkjörnungum, hafa þeir umfangsmikla, innri himnubundna sekki sem kallast skífur (himnuskífur). Blaðgræna er hluti af þessum himnum, ásamt mörgum próteinum ljóstillífunarinnar. Blágerlar hafa einnig peptíðóglýkanvegg og lípópólýsakkaríðlag sem tengist Gram-neikvæðum bakteríum.
Grænukorn af fyrsta stigs innra samlífisuppruna hafa skífur, hringlaga DNA litning og ríbósóm sem svipa til þeirra í blágerlum. Líkt og í hvatberum er hvert grænukorn umlukið tveimur himnum. Talið er að ytri himnan sé komin frá bólunni sem hýsillinn myndaði utan um samlífisveruna, og innri himnan sé komin frá frumuhimnu blágerilsins sem var innlimaður. Í hópi Archaeplastida sem kallast glaucophytar og í rhizarian-lífverunni Paulinella er þunnt peptíðóglýkanlag enn til staðar milli ytri og innri plastíðhimnanna. Öll önnur plastíð skortir þessar leifar af blágerlaveggnum.
Það eru einnig, líkt og í tilfelli hvatbera, sterkar vísbendingar um að mörg gen innri samlífislífverunnar hafi flust yfir í kjarnann. Plastíð, líkt og hvatberar, geta ekki lifað sjálfstætt utan hýsilsins. Auk þess, líkt og hvatberar, verða plastíð til við skiptingu annarra plastíða og eru aldrei byggð frá grunni. Rannsakendur hafa bent á að innra samlífisatburðurinn sem leiddi til Archaeplastida hafi átt sér stað fyrir 1 til 1,5 milljörðum ára, að minnsta kosti 500 milljónum ára eftir að steingervingasagan bendir til að heilkjörnungar hafi verið til staðar.
Ekki eru öll plastíð í heilkjörnungum komin beint af fyrsta stigs innra samlífi. Sumir af helstu hópum þörunga urðu ljóstillífandi við annars stigs innra samlífi, það er að segja með því að taka inn annaðhvort grænþörunga eða rauðþörunga (báðir frá Archaeplastida) sem innri samlífislífverur (Mynd 23.4). Fjölmargar smásjár- og erfðafræðirannsóknir hafa stutt þessa niðurstöðu. Annars stigs plastíð eru umlukin þremur eða fleiri himnum, og sum annars stigs plastíð hafa jafnvel kjarngerfil, greinilega leif af kjarna innri samlífisþörungsins. Það eru jafnvel tilfelli þar sem þriðja stigs eða hærra stigs innra samlífisatburðir eru bestu skýringarnar á eiginleikum sumra plastíða í heilkjörnungum.
Sjónræn tenging

Hvaða sannanir eru fyrir því að hvatberar hafi verið innlimaðir í forföður heilkjörnunga á undan grænukornum?
Mynd 23.5: Allar heilkjörnungafrumur hafa hvatbera eða afleiður þeirra, en ekki allar heilkjörnungafrumur hafa grænukorn.
Þróunartenging
Annars stigs innra samlífi í Chlorarachniophyta
Innra samlífi felur í sér að ein fruma gleypir aðra til að mynda, með tímanum, samþróað samband þar sem hvorug fruman gæti lifað ein. Grænukorn rauð- og grænþörunga eru til dæmis komin til vegna þess að forfaðir dreifkjörnunga gleypti ljóstillífandi blágeril.
Þessar vísbendingar benda til þess möguleika að forfaðir frumu (sem innihélt þegar ljóstillífandi innri samlífislífveru) hafi verið gleyptur af annarri heilkjörnungafrumu, sem leiddi til annars stigs innra samlífis. Sameinda- og formfræðilegar vísbendingar benda til þess að frumverur af hópi chlorarachniophyta séu komnar af annars stigs innra samlífisatburði. Chlorarachniophytar eru sjaldgæfir þörungar sem eru upprunnir í sandi hitabeltishafa. Þeir eru flokkaðir í Rhizaria-yfirhópinn. Chlorarachniophytar eru netlaga amöbur sem teygja út þunna umfrymisþræði sem tengja þær við aðra chlorarachniophyta í umfrymisneti. Talið er að þessar frumverur hafi orðið til þegar heilkjörnungur gleypti grænþörung, en sá síðarnefndi hafði áður stofnað til innra samlífis við ljóstillífandi blágeril (Mynd 23.6).
Nokkrar vísbendingar styðja að chlorarachniophytar hafi þróast við annars stigs innra samlífi. Grænukornin í grænþörungnum sem er innri samlífislífvera eru enn fær um ljóstillífun, þannig að chlorarachniophyta eru ljóstillífandi. Grænþörungurinn sem er innri samlífislífvera sýnir einnig kjarngerfil. Í raun virðist sem chlorarachniophytar séu afurð þróunarlega nýlegs annars stigs innra samlífisatburðar. Plastíð chlorarachniophyta eru umlukin fjórum himnum: Fyrstu tvær samsvara innri og ytri himnum ljóstillífandi blágerilsins, sú þriðja samsvarar frumuhimnu grænþörungsins og sú fjórða samsvarar bólunni sem umkringdi grænþörunginn þegar hann var gleyptur af forföður chlorarachniophyta. Í öðrum þróunarlínum sem fólust í annars stigs innra samlífi er aðeins hægt að greina þrjár himnur umhverfis plastíð. Þetta er nú túlkað sem stigvíst tap á himnu í gegnum þróunarsöguna.
Annars stigs innra samlífi er ekki einsdæmi fyrir chlorarachniophyta. Annars stigs plastíð finnast einnig í Excavata og Chromalveolata. Í Excavata leiddi annars stigs innra samlífi grænþörunga til eugleníða, en í Chromalveolata leiddi annars stigs innra samlífi rauðþörunga til þróunar plastíða í skoruþörungum, gródýrum og stramenopillum.