13.1 Litningakenningin og erfðatengsl
13.1 Litningakenningin og erfðatengsl
Í lok þessa hluta muntu geta gert eftirfarandi:
Löngu áður en vísindamenn sáu litninga í smásjá, hóf faðir nútímaerfðafræði, Gregor Mendel, að rannsaka erfðir árið 1843. Með bættri smásjártækni seint á 19. öld gátu frumulíffræðingar litað og skoðað frumulíffæri með litarefnum og fylgst með virkni þeirra við frumuskiptingu og meiósu. Við hverja mítósuskiptingu eftirmynduðust litningar, þéttust úr óreglulegum kjarnamassa í aðgreindar X-lagaðar byggingar (pör af eins systurlitningsþráðum) og færðust að aðskildum frumuskautum.
Litningakenning erfða
Sú tilgáta að litningar gætu verið lykillinn að skilningi á erfðum leiddi til þess að nokkrir vísindamenn skoðuðu ritverk Mendels og endurmatu líkan hans með tilliti til hegðunar litninga við mítósu og meiósu. Árið 1902 tók Theodor Boveri eftir því að eðlilegur fósturþroski ígulkers á sér ekki stað nema litningar séu til staðar. Sama ár tók Walter Sutton eftir aðskilnaði litninga í dótturfrumur við meiósu (Mynd 13.2). Saman leiddu þessar athuganir til litningakenningar erfða, sem skilgreindi litninga sem erfðaefnið sem bæri ábyrgð á mendelskum erfðum.

Litningakenning erfða samræmdist lögmálum Mendels, sem eftirfarandi athuganir studdu:
Þrátt fyrir skort á beinum sönnunum fyrir því að litningar bæru einkenni, leiddi hin sannfærandi fylgni milli hegðunar litninga við meiósu og óhlutbundinna lögmála Mendels til þess að vísindamenn settu fram litningakenningu erfða. Gagnrýnendur bentu á að einstaklingar hefðu mun fleiri einkenni sem erfðust óháð hvert öðru en þeir hefðu litninga. Um tíu árum eftir að kenningin var sett fram var Eleanor Carothers sú fyrsta til að uppgötva efnislegar sannanir sem studdu hana; hún sá óháða röðun litninga í engisprettum. Síðan, eftir nokkurra ára vinnu við víxlanir á ávaxtaflugunni, Drosophila melanogaster, lagði Thomas Hunt Morgan fram frekari tilraunagögn til stuðnings litningakenningu erfða.
Erfðatengsl og fjarlægðir
Vinna Mendels benti til þess að einkenni erfðust óháð hvert öðru. Morgan fann 1:1 samsvörun milli einkennis sem aðskildist og X-litningsins, sem benti til þess að handahófskenndur aðskilnaður litninga væri efnislegur grunnur líkans Mendels. Þetta sýndi einnig fram á að tengd gen raska spám Mendels. Að hver litningur geti borið mörg tengd gen skýrir hvernig einstaklingar geta haft mun fleiri einkenni en þeir hafa litninga. Hins vegar bentu rannsakendur á rannsóknarstofu Morgans á að genasamsætur staðsettar á sama litningi erfðust ekki alltaf saman. Við meiósu urðu tengd gen einhvern veginn ótengd.
Samstæð endurröðun
Árið 1909 sá Frans Janssen krossmót—staðinn þar sem litningsþræðir snertast og geta skipst á bútum—fyrir fyrstu meiósuskiptinguna. Hann lagði til að genasamsætur yrðu ótengdar og litningar skiptust efnislega á bútum. Þegar litningar þéttust og pöruðust við samstæður sínar, virtust þeir eiga samskipti á tilteknum stöðum. Janssen lagði til að þessir staðir samsvöruðu svæðum þar sem litningsbútar skiptust á. Við vitum nú að pörun og samspil milli samstæðra litninga, eða litningapörun, gerir meira en aðeins að skipuleggja samstæðurnar fyrir flutning í aðskildar dótturfrumur. Þegar þeir eru paraðir ganga samstæðir litningar í gegnum gagnkvæm efnisleg skipti á örmum sínum við samstæða endurröðun, eða einfaldlega litningavíxl.
Til að skilja betur þá tegund tilraunaniðurstaðna sem rannsakendur fengu á þessum tíma, skulum við íhuga arfblendinn einstakling sem erfði ríkjandi móður-genasamsætur fyrir tvö gen á sama litningi (eins og A og B) og tvær víkjandi föður-genasamsætur fyrir sömu gen (eins og a og b). Ef genin eru tengd mætti búast við að þessi einstaklingur myndi kynfrumur sem eru annaðhvort AB eða ab í hlutfallinu 1:1. Ef genin eru ótengd ætti einstaklingurinn að mynda AB, Ab, aB og ab kynfrumur með jafnri tíðni, samkvæmt hugmynd Mendels um óháða röðun. Þar sem þær samsvara nýjum samsetningum genasamsæta, eru arfgerðirnar Ab og aB endurraðaðar gerðir sem verða til við samstæða endurröðun í meiósu. Foreldragerðir eru afkvæmi sem sýna sömu samsetningu genasamsæta og foreldrar þeirra. Morgan og samstarfsmenn hans fundu hins vegar að þegar þeir prófvíxluðu slíkum arfblendnum einstaklingum við arfhreint víkjandi foreldri (AaBb aabb), komu bæði foreldragerðir og endurraðaðar gerðir fram. Til dæmis gætu 950 afkvæmi fundist sem væru annaðhvort AaBb eða aabb, en einnig 50 afkvæmi sem væru annaðhvort Aabb eða aaBb. Þessar niðurstöður bentu til þess að tengsl ættu sér oftast stað, en marktækur minnihluti afkvæma væri afurð endurröðunar.
Ein af tilraununum á rannsóknarstofu Morgans, sem fól í sér víxlanir á flugum fyrir tvö einkenni, líkamslit (grár eða svartur) og vænglögun (venjulegir og rýrnaðir), sýndi fram á endurröðunarviðburði sem leiða til þróunar á svipgerðum sem eru frábrugðnar foreldrum (Mynd 13.3).
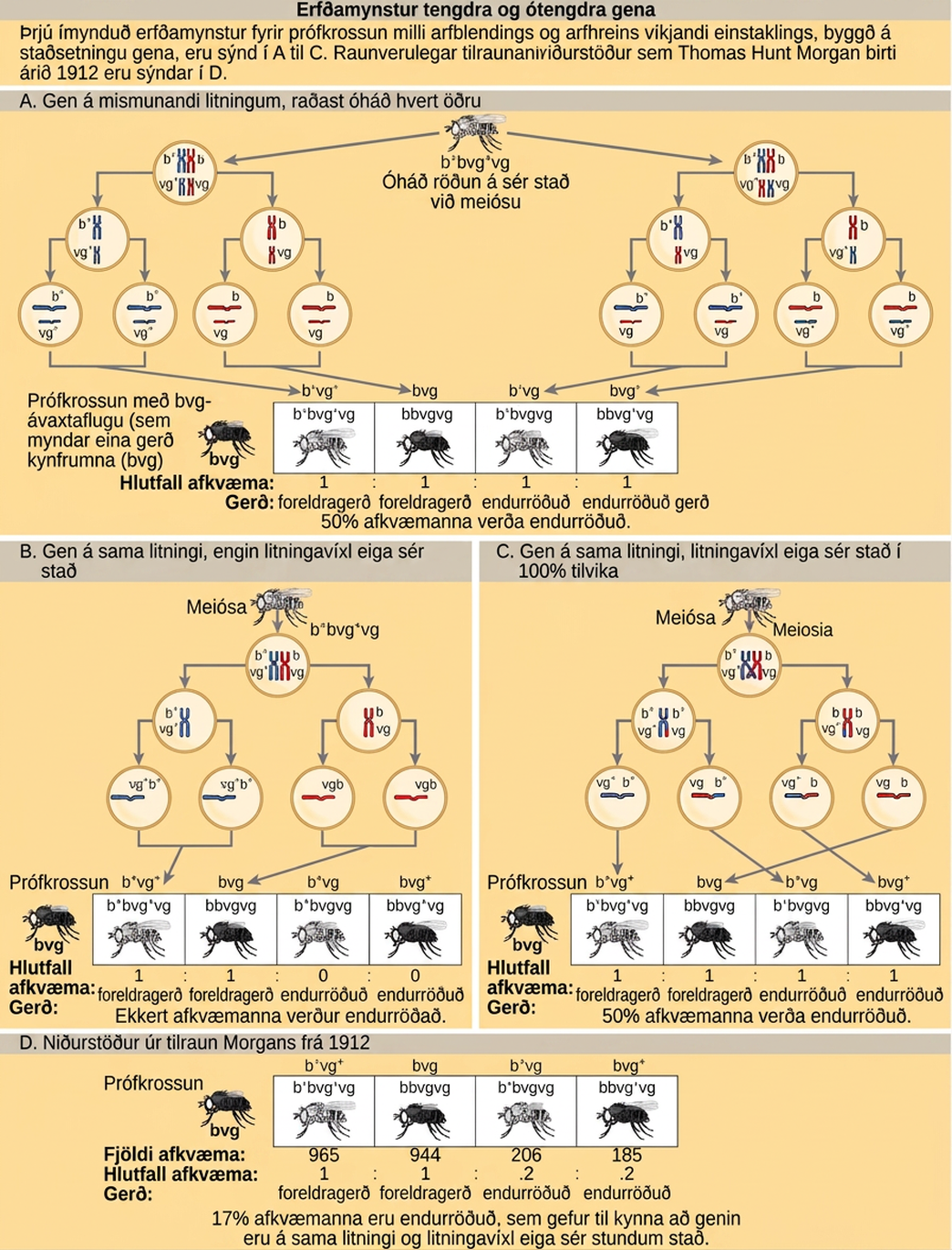
Í prófvíxlun fyrir tvö einkenni eins og þá sem hér er sýnd, getur spáð tíðni endurraðaðra afkvæma verið 60 prósent? Hvers vegna eða hvers vegna ekki?
Genakort
Janssen hafði ekki tæknina til að sýna fram á litningavíxl svo það var áfram óhlutbundin hugmynd sem vísindamenn trúðu almennt ekki. Vísindamenn töldu krossmót vera tilbrigði við litningapörun og gátu ekki skilið hvernig litningar gætu brotnað og tengst aftur. Samt voru gögnin skýr um að tengsl áttu sér ekki alltaf stað. Að lokum þurfti ungan grunnnema og eina „vökunótt“ til að skýra stærðfræðilega vandamálið um tengsl og endurröðun.
Árið 1913 safnaði Alfred Sturtevant, nemandi á rannsóknarstofu Morgans, niðurstöðum frá rannsakendum á rannsóknarstofunni og tók þær með sér heim eitt kvöldið til að velta þeim fyrir sér. Morguninn eftir hafði hann búið til fyrsta „litningakortið“, línulega framsetningu á röð gena og hlutfallslegri fjarlægð á litningi (Mynd 13.4).

Hver af eftirfarandi fullyrðingum er sönn?
Eins og Mynd 13.4 sýnir, með því að nota tíðni endurröðunar til að spá fyrir um erfðafjarlægð, getum við ályktað um hlutfallslega röð gena á litningi 2. Gildin tákna kortafjarlægðir í sentímorgan (cM), sem samsvara tíðni endurröðunar (í prósentum). Þess vegna voru genin fyrir líkamslit og vængstærð með 65,5 − 48,5 = 17 cM millibili, sem gefur til kynna að móður- og föður-genasamsætur fyrir þessi gen endurraðist í 17 prósentum afkvæma, að meðaltali.
Til að búa til litningakort gerði Sturtevant ráð fyrir að genum væri raðað í röð á þráðlaga litninga. Hann gerði einnig ráð fyrir að tíðni endurröðunar milli tveggja samstæðra litninga gæti átt sér stað með jöfnum líkum hvar sem er eftir endilöngum litningnum. Með þessar forsendur að leiðarljósi setti Sturtevant fram þá tilgátu að genasamsætur sem væru langt hver frá annarri á litningi væru líklegri til að skiljast að við meiósu einfaldlega vegna þess að svæðið þar sem endurröðun gæti átt sér stað væri stærra. Aftur á móti væru genasamsætur sem væru nálægt hver annarri á litningnum líklegar til að erfast saman. Meðalfjöldi víxlana milli tveggja genasamsæta — það er að segja tíðni endurröðunar þeirra — fylgdi erfðafjarlægð þeirra hvor frá annarri, miðað við staðsetningu annarra gena á þeim litningi. Ef litið er til dæmisins um víxlun milli AaBb og aabb hér að ofan, gætum við reiknað tíðni endurröðunar sem 50/1000 = 0,05. Það er að segja, líkurnar á víxlun milli gena A/a og B/b voru 0,05, eða 5 prósent. Slík niðurstaða myndi gefa til kynna að genin væru örugglega tengd, en að þau væru nógu langt í sundur til að víxlun gæti átt sér stað stöku sinnum. Sturtevant skipti erfðakorti sínu í kortaeiningar, eða sentímorgan (cM), þar sem 0,01 tíðni endurröðunar samsvarar 1 cM.
Með því að setja genasamsætur upp á línulegu korti, gaf Sturtevant til kynna að gen geti verið allt frá því að vera algerlega tengd (tíðni endurröðunar = 0) yfir í að vera algerlega ótengd (tíðni endurröðunar = 0,5) þegar gen eru á mismunandi litningum eða mjög langt í sundur á sama litningi. Algerlega ótengd gen samsvara þeirri tíðni sem Mendel spáði fyrir um fyrir óháða samröðun í tvíblendingsvíxlun. Tíðni endurröðunar upp á 0,5 gefur til kynna að 50 prósent afkvæma séu endurraðaðir einstaklingar og hin 50 prósentin séu af foreldragerðum. Það er að segja, hver tegund samsetningar genasamsæta kemur fyrir með jafnri tíðni. Þessi framsetning gerði Sturtevant kleift að leggja saman fjarlægðir milli nokkurra gena á sama litningi. Hins vegar, þegar erfðafjarlægðir nálguðust 0,50, urðu spár hans ónákvæmari því það var ekki ljóst hvort genin væru mjög langt í sundur á sama litningi eða á mismunandi litningum.
Árið 1931 sýndu Barbara McClintock og Harriet Creighton fram á víxlun samstæðra litninga í maísplöntum. Nokkrum vikum síðar sýndi Curt Stern fram á samstæða endurröðun í Drosophila með smásjárskoðun. Stern tók eftir nokkrum X-tengdum svipgerðum sem tengdust byggingarlega óvenjulegu og ósamstæðu X-litningapari þar sem annan X-litninginn vantaði lítinn endahluta, og hinn X-litningurinn var runninn saman við bút af Y-litningnum. Með því að víxla flugum, fylgjast með afkvæmum þeirra og skoða síðan litninga afkvæmanna, sýndi Stern fram á að í hvert sinn sem samsetning genasamsæta afkvæmisins vék frá annarri hvorri foreldrasamsetningunni, átti sér stað samsvarandi skipti á hluta af X-litningi. Notkun stökkbreyttra flugna með byggingarlega ólíka X-litninga var lykillinn að því að sjá afurðir endurröðunar því DNA raðgreining og önnur sameindatól voru ekki enn fáanleg. Við vitum nú að samstæðir litningar skiptast reglulega á hlutum við meiósu með því að rjúfa og tengja aftur DNA sína á nákvæmum stöðum. Aurora Ruiz-Herrera rannsakar til dæmis erfðafræðilega brotstaði á stöðum í litningum sem kallast brothætt svæði. Með því að bera kennsl á brothætt svæði í litningum sem menn og aðrir prímatar eiga sameiginleg, hefur Ruiz-Herrera veitt dýpri skilning á þróun spendýra og sérstaklega þróun manna.
Skoðaðu ferli Sturtevants við að búa til erfðakort á grundvelli tíðni endurröðunar hér.
Kortlögð einkenni Mendels
Samstæð endurröðun er algengt erfðafræðilegt ferli, en samt varð Mendel aldrei var við það. Hefði hann rannsakað bæði tengd og ótengd gen, hefði verið mun erfiðara fyrir hann að búa til samræmt líkan af gögnum sínum á grundvelli líkindaútreikninga. Vísindamenn sem hafa síðan kortlagt einkennin sjö sem Mendel rannsakaði á sjö litninga erfðamengis garðertunnar hafa staðfest að öll genin sem hann skoðaði eru annaðhvort á aðskildum litningum eða eru nægilega langt í sundur til að vera tölfræðilega ótengd. Sumir hafa haldið því fram að Mendel hafi verið gríðarlega heppinn að velja aðeins ótengd gen; á meðan aðrir spyrja sig hvort Mendel hafi hent gögnum sem bentu til tengsla. Hvað sem því líður, sá Mendel stöðugt óháða röðun vegna þess að hann rannsakaði gen sem voru í raun ótengd.